
Eusthenopteron foordi. Artwork by Nobu Tamura, 2007. Wikipedia, GNU Free Documentation/Creative Commons Attribution license.
| Sarcopterygii | ||
| The Vertebrates | Eusthenopteron |
| Vertebrates Home | Vertebrate | Vertebrate |
|
Abbreviated Dendrogram
Teleostomi
├─Neopterygii
└─Sarcopterygii
├─┬─Onychodontiformes
│ └─Actinistia
└─Rhipidistia
├─Dipnomorpha
└─Tetrapodomorpha
├─Rhizodontiformes
└─Osteolepiformes
├─Tristichopteridae
│ ├─Eusthenopteron
│ └─┬─Cabonnichthys
│ └─Mandageria
└─Elpistostegalia
├─Panderichthys
└─Tetrapoda
|
Contents
Overview |
Eusthenopteron foordi. Artwork by Nobu Tamura, 2007. Wikipedia, GNU Free Documentation/Creative Commons Attribution license. |
What can we say about Eusthenopteron that has not already been said, much better, by someone else? Eusthenopteron is the gold standard for osteolepiforms, the Morganucodon of sarcopterygians, and, in fact, probably the best known Paleozoic vertebrate of any taxon, bar none. Certainly it is the point of reference for all comparative studies in this part of phylospace, including our own. Eusthenopteron remains are known by the hundred from the Frasnian Escuminac Formation of Quebec, Canada. Erik Jarvik, were he remembered for nothing else, would still be guaranteed paleontological sainthood for the decades he spend grinding, millimeter by millimeter, through impeccably preserved specimens of Eusthenopteron and constructing detailed, three-dimensional models of its skull. Andrews & Westoll (1970) followed with a 120-page monograph on its post-cranial anatomy. Others have altered the terminology, and reinterpreted a few of the conclusions reached by Jarvik, Andrews and Westoll. No one takes issue with their observations. We decline to accept the pointless challenge of doing so.
However, the time has come to address that part of rhipidistian anatomy which first brought this group to the world's startled attention in the Nineteenth Century and has continued to raise eyebrows since -- the pectoral limb. For purposes of this section, we will confine ourselves to Eusthenopteron.
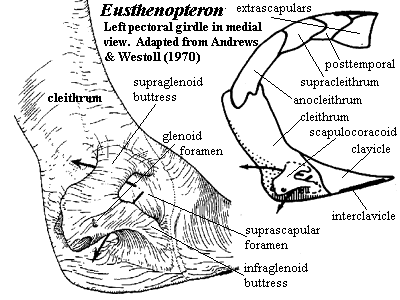
As with virtually every other sort of jawed fish, rhipidistians have the forelimb girdle stapled onto the back of the skull. In rhipidistians, this is accomplished by a chain of dermal bones beginning dorsally) with the median dorsal extrascapular, and running, to each side, through the lateral extrascapular, posttemporal, supracleithrum, and anocleithrum to the cleithrum, the largest element of the series. On the medial, or posteromedial, face of the cleithrum is a complex endochondral bone, the scapulocoracoid, through which the limb articulates. The limb girdle then continues ventrally through the clavicle and completes the circle with a median ventral interclavicle.
The cleithrum, with its long dorsal stem, brings to mind the amniote scapula. The similarity is not entirely fortuitous, since both scapula and cleithrum act to stabilize the forelimb against the body wall, while providing a degree of flexibility. However, the cleithrum of Eusthenopteron is a somewhat busier structure. It was anteriorly placed and tightly associated with the skull. Its anterior margin was overlapped by the opercular bones. On this unornamented anterior surface, the cleithrum probably anchored jaw muscles and the principle membranes covering the gills. The clavicle in Eusthenopteron was less elaborate than in rhizodonts and probably served a primarily structural role. Rather than attaching large adductor muscles, it merely anchored the limbs ventrally and, through an anterior process, may have provided support for the basibranchium and perhaps the lower jaw. The interclavicle is not known with certainty and was probably a small, flexible cartilaginous link.
The scapulocoracoid is relatively small. The glenoid fossa (see image below) faces posteroventrally and somewhat laterally. [2] The glenoid has a long extension which forms a direct articulation with the cleithrum. This is one of three legs of the tripodal scapulocoracoid. The other two legs are the more dorsal supraglenoid buttress and the more ventral infraglenoid buttress. The arched gap between the buttresses, directly opposite the glenoid, is the suprascapular foramen. The arched gaps above and below the glenoid are the supraglenoid and supracoracoid foramina, respectively. The glenoid fossa itself is somewhat pear-shaped, with the small end pointing toward the cleithrum. The articulating surface is slightly torted, forming a shape believed to be homologous with the "screw-shaped" glenoid of early tetrapods. Exactly how this assisted the fish is not known. Presumably it (a) adjusted the attitude of the fin at the end of a recovery stroke to prepare for a power stroke or (b) vice-versa, or it locked the fin in position for (c) braking, (d) turning, or (e) lifting the body against gravity. From its position, we would favor some combination of (a), (c) and (d); but the matter is speculative.
| Mesomere #1 | Humerus |
| Radial #1 | Radius |
| Postaxial Process #1 | Entepicondyle |
| Mesomere #2 | Ulna |
| Radial #2 | Intermedium |
| Mesomere #3 | Ulnare |
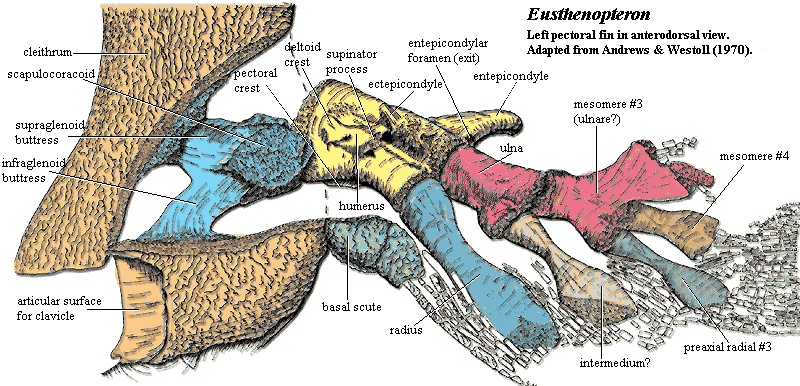
The limb itself is uniserial and composed of 4, perhaps 5, mesomeres. Each mesomere bears a an unbranched radial on its preaxial (more or less anterior) side. Mesomeres 1 and 3 -- presumably also mesomere 5 if it existed -- also bear a stout postaxial process. The homologies on which nearly everyone agrees are listed in the table in bold. More controversial assignments are not bolded. For an explanation of humeral structures, see the Humerus.
The humerus bears a number of structures that make its assignment indisputable. As mentioned it has a very large entepicondyle, and an entepicondylar foramen is also present. The ectepicondyle is also present, although it is located in an unconventionally proximal and median position. Many of the other lumps and bumps of the tetrapod humerus are present as well. The supinator process, for example, is conventionally placed with respect to the ectepicondyle, but similarly displaced from the usual tetrapod position. One unique feature of Eusthenopteron is a process which looks like an epicondyle projecting ventrally from the ventral face of the distal humerus, which Andrews & Westoll (1970) refer to as the "humeral process." It is not present in tetrapods or even in other tristichopterids.
 The limb bones are flattened distally and bear jointed lepidotrichia. The net effect is to create a dense fringe of lepidotrichia on the preaxial margin of the fin, extending well over onto the postaxial margin. In life, the scales of the limb overlapped the proximal portion of the lepidotrichia, creating the familiar lobe-finned appearance of sarcopterygians. The lepidotrichia are thin, dermal structures with no possible homology to the phalanges of the tetrapod digit. Likewise -- despite numerous attempts over the years -- no possible manipulation of the limb bones can create tetrapod digits. In living tetrapods, the digits are formed uniquely by a "digital arch" of cartilage across the distal limb. The digits develop as radial pulses of bone formation from near the center of the arch, accompanied by a variable amount of apoptosis (programmed cell death) between the developing phalanges. This radial pattern is quite distinct from the branching pattern of mesomere formation observed here. No trace of radial development appears in Eusthenopteron, although something quite similar to a digital arch must account for the pectoral limb of rhizodonts.
The limb bones are flattened distally and bear jointed lepidotrichia. The net effect is to create a dense fringe of lepidotrichia on the preaxial margin of the fin, extending well over onto the postaxial margin. In life, the scales of the limb overlapped the proximal portion of the lepidotrichia, creating the familiar lobe-finned appearance of sarcopterygians. The lepidotrichia are thin, dermal structures with no possible homology to the phalanges of the tetrapod digit. Likewise -- despite numerous attempts over the years -- no possible manipulation of the limb bones can create tetrapod digits. In living tetrapods, the digits are formed uniquely by a "digital arch" of cartilage across the distal limb. The digits develop as radial pulses of bone formation from near the center of the arch, accompanied by a variable amount of apoptosis (programmed cell death) between the developing phalanges. This radial pattern is quite distinct from the branching pattern of mesomere formation observed here. No trace of radial development appears in Eusthenopteron, although something quite similar to a digital arch must account for the pectoral limb of rhizodonts.
Creationists frequently speak disparagingly of the absence of a "fish with legs." But Eusthenopteron and its relatives are unquestionably fish with legs. The interesting part is that they have no hands or feet. Rhizodonts, which seem to be off the main line of tetrapod evolution, have both limbs and hands. So the "pod" of the tetrapods is not a new thing. The question which needs answering is, where did this elegant little developmental program come from? At the moment, we don't know. ATW031105.
[2] Note that the glenoid fossa is the cavity in which the humerus articulates. The glenoid foramen is a small canal located on the side of the glenoid opposite the glenoid fossa.
Image: Life reconstruction from Videoscapes.
 Eusthenopteron: E. foordi Whiteaves, 1881.
Eusthenopteron: E. foordi Whiteaves, 1881.
Range: Late Devonian (Frasnian) of North America (Canada).
Phylogeny: Tristichopteridae: (Cabonnichthys + Mandageria) + *.
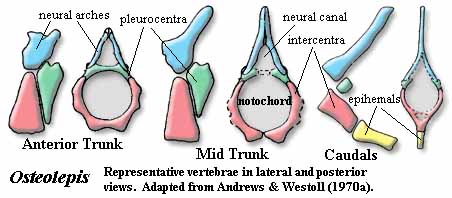
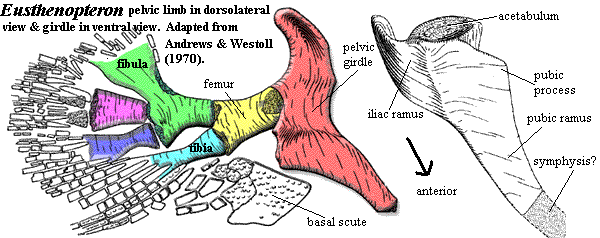
Characters: up to 1.5 m [AW70]; postspiracular located between lateral extrascapular and opercular, but also extends anteriorly between tabular & squamosal [JA97]; nasolacrimal duct present; choanae present; parasymphysial fang pair absent [JA97]; extensive coronoid marginal dentition [JA97]; body with somewhat triangular cross-section with large, flat ventral surface [AW70]; vertebrae rachitomous, generally similar to osteolepidids (image) [AW70]; pleurocentra only associated with base of next anterior neural arch [AW70a]; pleurocentra always paired [C88]; pleurocentra small, rounded, irregular & unfinished, lying posterior to other elements [AW70]; intercentra paired in trunk, but fused anteriorly and caudally [C88]; dorsal parts of intercentra with unfinished bone [AW70]; ventral end of intercentra notched posteriorly for passage of intersegmental arteries [AW70]; intercentra continuous with hemal arch in tail [C88]; central elements separated by cartilage from neural arches [C88]; neural arches gracile, joined distally, with expanded areas for nerve cord and dorsal ligament [AW70]; arches with smooth periosteal bone externally, unfinished (probably lines with cartilage) internally [AW70]; "incipient zygapophyses" at the level of the top of the neural canal, with matching variations in facing vertebrae [AW70]; neural arches do not overlap intercentra [AW70]; small ribs on mid-trunk vertebrae with capitulum articulating with parapophysis at mid-height on intercentrum, with dorsal end (tuberculum) unfinished, and with a minimal shaft directed dorsolaterally & posteriorly [AW70]; sacral vertebra with greatly enlarged parapophyses, oriented ventrolaterally, probably was attachment for soft-tissue connection with pelvic girdle [AW70]; caudal vertebrae with intercentra short, hemal spines present, pleurocentra small or absent & ribs absent [AW70]; all fins except caudal with large basal scute [AW70]; 1st dorsal fin supported by single large, thick triangular basal with irregular ventral base & bearing single radial [AW70]; anal & 2nd dorsal very similar, with single, quadrangular basal articulating with neural element (1 neural spine or 2 hemal spines) and bearing 3 radials which are waisted at midlength [AW70]; anal and second dorsal almost opposite each other, with anal slightly more posterior [AW70]; 2nd dorsal radials may be separated by "steps" [AW70]; anal fin basal with rod-like proximal section [AW70a]; long caudal peduncle [JA97]; caudal (and other) fins with relatively high aspect ratio [JA97]; caudal fin triphiceral [AW70]; paired fins at ventrolateral angle of body [AW70]; shoulder girdle extends far anteriorly & provides attachment for jaw muscles [AW70]; for shape of cleithrum, see image [AW70]; anterior edge of cleithrum runs under operculum and has smooth area for attachment of gill membrane [AW70]; clavicle with anterior muscle attachment area (compare Rhizodus) [AW70a]; clavicle dorsal process without internally hollowed region [AW70a]; interclavicle very small [AW70]; scapulocoracoid single ossification with 3 articulations on cleithrum [AW70]; glenoid fossa piriform & twisted similar to screw-shape glenoid of early tetrapods [AW70]; glenoid oriented mostly posterior, braced by supra- and infraglenoid buttresses [AW70]; buttresses form supraglenoid and supracoracoid foramina & anterior suprascapular foramen [AW70]; approximately 4 mesomeres [C+02]; no radials on distal radius or postaxially [C+02]; scales irregularly circular ornamented with ridges & elongate tubercles [AW70]; scales with internal drop-shaped boss [AW70]; fin scales small [AW70]; pectoral preaxial edge reinforced with thick ridge scales & lepidotrichia [AW70]; preaxial, proximal scales large, hemicylindrical, with atypical ornament [AW70]; large basal scute dorsal [2] to fin with long median ridge to attach to body [AW70];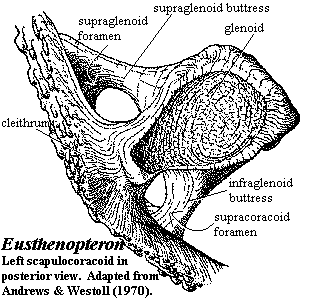 marginal lepidotrichia from mid-radius, around distal end to distal end of ulna [AW70]; pectoral fin terminal bones are flattened distally [AW70]; pectoral fin bones as in image [AW70] [3]; humerus as flattened cylinder, dorsal surface with deltoid & supinator processes [4] and ectepicondyle [AW70]; humeral head piriform and convex, with shallow groove [AW70]; deltoid & supinator processes sometimes fused [AW70]; latissimus dorsi process absent [AW70]; entepicondyle very specialized by comparison to generalized postaxial processes [AW70]; humerus, ventral surface with strong ridge continuous with entepicondyle proximally & running transversely & proximally to pectoral process [AW70]; second humeral process on ventral face between entepicondyle & ulnar articulation ~half as large as entepicondyle and also connected to ventral ridge [5] [AW70]; ulnar articulation slightly distal to radial [AW70]; both ulnar & radial articulations with a small ventral articular surface in addition to distal articulation [AW70]; note unexpected absence of postaxial process on mesomere #2 (ulna) [AW70]; ulna stout & half length of radius [AW70]; ulnar distal articulations flat [AW70]; all more distal components are much flattened [AW70]; radius also thin & bladelike distally [AW70]; pelvic limb similar structure, but smaller -- see image [AW70]; pelvic girdle with single ossification of pubis + ileum [AW70]; acetabulum on buttress, facing posterolaterally [AW70]; acetabulum with small dorsal overhang (supraacetabular shelf) [AW70]; pubic ramus triangular in cross-section [AW70]; iliac blade with groove on medial face above acetabulum for muscle attachment [AW70]; acetabulum concave, fitting closely to femoral head [AW70]; possible attachment to sacral vertebra [AW70]; ischial ramus and obturator foramen absent [AW70] (contra [L+01] following Jarvik's terminology); femur relatively featureless, with groove dorsally between distal articulations [AW70]; femur ventral face concave, probably accommodating the PIFE & homologous to intertrochanteric fossa [AW70] [6]; femur distal articulations relatively flat [AW70]; preaxial radials are distally flattened as on pectoral limb [AW70].
marginal lepidotrichia from mid-radius, around distal end to distal end of ulna [AW70]; pectoral fin terminal bones are flattened distally [AW70]; pectoral fin bones as in image [AW70] [3]; humerus as flattened cylinder, dorsal surface with deltoid & supinator processes [4] and ectepicondyle [AW70]; humeral head piriform and convex, with shallow groove [AW70]; deltoid & supinator processes sometimes fused [AW70]; latissimus dorsi process absent [AW70]; entepicondyle very specialized by comparison to generalized postaxial processes [AW70]; humerus, ventral surface with strong ridge continuous with entepicondyle proximally & running transversely & proximally to pectoral process [AW70]; second humeral process on ventral face between entepicondyle & ulnar articulation ~half as large as entepicondyle and also connected to ventral ridge [5] [AW70]; ulnar articulation slightly distal to radial [AW70]; both ulnar & radial articulations with a small ventral articular surface in addition to distal articulation [AW70]; note unexpected absence of postaxial process on mesomere #2 (ulna) [AW70]; ulna stout & half length of radius [AW70]; ulnar distal articulations flat [AW70]; all more distal components are much flattened [AW70]; radius also thin & bladelike distally [AW70]; pelvic limb similar structure, but smaller -- see image [AW70]; pelvic girdle with single ossification of pubis + ileum [AW70]; acetabulum on buttress, facing posterolaterally [AW70]; acetabulum with small dorsal overhang (supraacetabular shelf) [AW70]; pubic ramus triangular in cross-section [AW70]; iliac blade with groove on medial face above acetabulum for muscle attachment [AW70]; acetabulum concave, fitting closely to femoral head [AW70]; possible attachment to sacral vertebra [AW70]; ischial ramus and obturator foramen absent [AW70] (contra [L+01] following Jarvik's terminology); femur relatively featureless, with groove dorsally between distal articulations [AW70]; femur ventral face concave, probably accommodating the PIFE & homologous to intertrochanteric fossa [AW70] [6]; femur distal articulations relatively flat [AW70]; preaxial radials are distally flattened as on pectoral limb [AW70].
Note: [1] "In Eusthenopteron (a representative osteolepiform) and the tetrapods, each nasal cavity consists of a junction of three ducts surrounded by bone) which mutually connect the external naris (homologous with the anterior naris of most fish), orbit (eye socket), and internal naris choana, which opens into the mouth cavity en route to the trachea). The connection to the orbit, termed the nasolacrimal duct, conducts excessive tear fluid from the eyes into the nasal cavity (hence crying leads to a runny nose). Its presence in Eusthenopteron suggests that orbital fluid allowed the eye to be exposed to the air for extended periods without desiccation. The small size of the external naris in Eusthenopteron suggests that the primary adaptive benefit of the choana was improved smelling capability; most respiration was would still have been conducted through the mouth." Fall'96Syllabus. [2] In Megalichthys & Osteolepis the basal scutes are found both dorsal & ventral to the pectoral fins [AW70]. [3] Andrews & Westoll [AW70] reconstruct the fin as having projected posteroventrally, with the dorsal face tilted to face somewhat laterally. They speculate that, at rest, the postaxial edge was accommodated in an embayment of the body wall. [4] what does a supinator process do in an animal with no wrist? Note also that Coates [Co96] disagrees with the [AW70] interpretation to some extent and asserts that the structure identified by [AW70] as the deltoid process is actually the latissimus dorsi process. [5] Andrews & Westoll [AW70] suppose that this process served as a sort of doorstop to prevent the ulna from swinging out postaxially. [6] Andrews & Westoll [AW70] note that the PIFE would have drawn the fin ventromedially and ?anteriorly for braking.
Image: For cranial bones, see also image at Rhipidistia.
Links: Cross-section of a Eusthenopteron- labyrinthodont tooth; Eusthenopteron エウステノプテロン; Kvastfinnefisk; Evolution of Limbs from the Fossil Record and Molecular Biology; Fall'96Syllabus; Vertebral development in the Devonian sarcopterygian fish ... abstract); Geology 143; The evolution of tetrapod limbs: How did fishes obtain their limbs?; BSC 310 LECTURE 11.; Comparative Vertebrate Anatomy - Lecture Notes 5; Eusthenopteron (cast of fin); Past lives- Chronicles of Canadian Paleontology (history of discovery. Also in French); Devonian Times - Tetrapods Answer one of the premier web sites on the general area); Musée McCord Museum - "Eusthenopteron Foorai" fossil, from ... so they can't spell! It's still a very nice image); Scientific American- Getting a Leg Up on Land 2005 article by Prof. Clack).
References: Andrews & Westoll (1970) [AW70], Andrews & Westoll (1970a) [AW70a], Carroll 1988) [C88], Coates (1996) [Co96]; Coates et al. 2002), Johanson & Ahlberg (1997) [JA97]; Liem et al. (2001) [L+01]. ATW060125.
checked ATW051025
{kind=link}
{kind=link}
{kind=link}