
Hovasaurus boulei, a tangasaurid from the Late Permian of Madagascar. Illustration by Nobu Tamura (Wikipedia)
| Diapsida | ||
| The Vertebrates | Younginiformes |
| Vertebrates Home | Vertebrate | Vertebrate |
|
Abbreviated Dendrogram
Amniota
├─Synapsida
└─Sauropsida
├─Anapsida
╘═Eureptilia
│
└─Diapsida
╞═Araeoscelida
└─Neodiapsida
├─Youngina Younginiformes
├─Thadeosaurus
└─┬─Tangasaurinae
└─┬─Claudiosaurus
├─Weigeltisauridae
└─┬─┬─Thalattosauria
│ └─Ichthyosauria
├?─Hupehsuchia
├?─Omphalosaurus
├?─Saurosphargidae
└─Sauria
├?─Choristodera
├─Lepidosauromorpha
└─Archosauromorpha |
Contents
Overview |
Hovasaurus boulei, a tangasaurid from the Late Permian of Madagascar. Illustration by Nobu Tamura (Wikipedia) |
The Younginiformes began life as the Eosuchia, a group (of ordinal rank in Linnaean taxonomy) coined by Broom in 1914 to refer to early and generalised diapsid reptiles, on the basis of Youngina,known from the late Permian of South Africa. The group eventually became a wastebasket taxon for any early generalised diapsids that are not obviously lepidosaurs or archosaurs. As a result, Romer suggested the order Younginiformes to be applied strictly to those forms of primitive diapsid nature. Romer was dissatisfied with this and coined the name Younginiformes is a replacement
With the rise of Hennig-based cladistic methdology, Benton 1985, Gauthier et al, and Evans 1988 placed them in the lepidosauromorphs, as the sister taxon to Lepidosauria. Laurin 1991 moved them to the base of Neodiapsida, and these findings have been accepted by other workers; see Diapsid phylogeny for more. The following synthetic consensus view, based on Carroll, 1988, Gower, 1996, Frey, Sues & Munk, 1997, and Gao & Fox, 1998, is from Mikko's Phylogeny Archive. It doesn't represent a particular cladogram, but rather a dendrogram mash-up of different Evolutionary Systematic and Cladistic sources:
|
■──○ Neodiapsida Gauthier, 1984 sensu Benton 1985
├?─○†Weigeltisauridae Piveteau 1926 (†Coelurosauravus Piveteau 1926)
└─┬─†Apsisaurus witteri Laurin 1991 (now known to be a varanopsid pelycosaur!)
└─○Eosuchia Broom, 1924 sensu deBraga & Rieppel, 1997
╞?═○†Eosuchia Broom, 1924 [Younginiformes Romer, 1945]
│ ├?─†Acerosodontosaurus piveteaui Currie, 1980 [Acerosodontosauridae]
│ ├?─†Noteosuchus colletti (Watson, 1912) Broom, 1925 [Eosuchus colletti Watson, 1912]
│ ├─†Galesphyrus capensis Broom 1914 [Galesphyridae Currie 1981]
│ ├─○†Younginidae Broom, 1914
│ │ ├─†Heleosuchus griesbachi Broom 1913
│ │ ├─†Thadeosaurus colcanapi Carroll, 1981
│ │ └─†Youngina capensis Broom, 1914 [Youngoides romeri, Youngopsis]
│ └─○†Tangasauridae Haughton, 1924
│ ├─†Hovasaurus boulei Currie, 1981
│ ├─†Tangasaurus mennelli Currie, 1982
│ └─†Kenyasaurus mariakanensis Harris & Carroll, 1977
└─┬?─†Claudiosaurus germaini Carroll, 1981 [Claudiosauridae Carroll, 1981 ]
├?─┬─○†Lazarussuchus Hecht, 1992; U.Olig.-L.Mioc. Eu. [Lazarussaurus (lapsus calami)]
│ │ ├─†L. inexpectatus Hecht, 1992
│ │ └─†L. dvoraci Evans & Klembara, 2005
│ └?─†Choristodera
├?─†"Ichthyosauromorpha"
└─Sauria Gauthier, 1984
|
In recent years the hand-coded Hennigian analysis has been replaced by computer-based statistical parsimony analysis using large numbers of traits from a larger range of taxa. These result in hundreds of trees, from which the most parsimonious (the ones of shortest length) are selected. This different methodology results in very different tree topology (the shape of the cladogram; i.e. the arrangement of the nodes and taxa). When applied to the Younginiformes this resulted in a very different, and much less tidy, arrangement. The following abstract is from Bickelmann, Müller & Reisz, 2009:
"A restudy of the Upper Permian diapsid Acerosodontosaurus piveteaui from Madagascar indicates that the bone formerly identified as the quadratojugal is a fragment of a rib. This in turn implies that, in contrast to previous studies, the lower temporal arcade must be considered incomplete and derived relative to the ancestral condition. Since the phylogenetic position of Acerosodontosaurus is poorly understood, the taxon was entered into a modified phylogenetic data matrix of diapsid reptiles, and the purported monophyly of "Younginiformes" was tested for the first time by including all potential members of the clade as separate taxa, as well as other taxa from the same deposits. The results of the phylogenetic analysis do not support the monophyly of "younginiform" reptiles. Instead, most taxa cluster unresolved at the base of Neodiapsida, a finding that has important implications for the understanding of early diapsid evolution because it suggests that early neodiapsids represent several distinct evolutionary lineages. Acerosodontosaurus and Hovasaurus do form a clade, a finding consistent with the stratigraphic age and biogeography of these taxa."
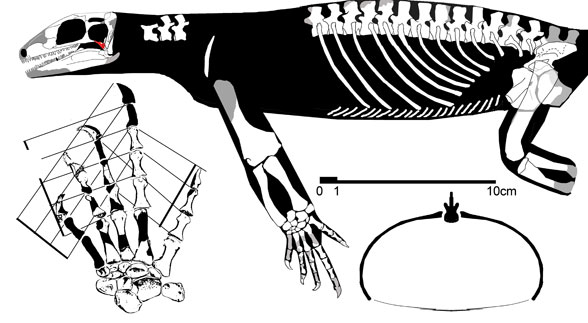 Illustration of Acerosodontosaurus piveteaui, by David Peters, Reptile Evolution. The quadratojugal bone, shown in red, is determined by Bickelmann et al. 2009 to be a portion of rib. This leads to a revolution in early diapsid relationships, including demolishing the monophyletic Younginiforms (Peters himself rejects this interpretation). |
The resulting cladogram is represented as follows (from Mikko's Phylogeny Archive):
|
■──○Neodiapsida
├─†Youngina
├─†Kenyasaurus
├─†Lanthanolania
├─†Thadeosaurus
├─†Galesphyrus
└─┬─┬─†Tangasaurus
│ └─┬─†Hovasaurus
│ └─†Acerosodontosaurus
└─┬─†Claudiosaurus
└─┬─┬─†Palaeagama
│ └─†Saurosternon
└─┬─†Coelurosauravus
└─○?Sauria
├─┬─†Thalattosauriformes
│ └─†"Ichthyosauromorpha"
└─┬─○Lepidosauromorpha
│ ├─†Choristodera
│ └─┬─†Sauropterygia
│ └─┬─Testudinata
│ └─┬─†Kuehneosauridae
│ └─Lepidosauriformes
└─○Archosauromorpha
╞═†"Protorosauria"
└─┬─†Trilophosauria
└─┬─†Rhynchosauria
└─Archosauriformes
|
In contrast to the earlier work of Bentan, Evans, and Gauthier et al, the Younginiformes have become an assemblage of basal Neodiapsids, and Claudiosaurus and Coelurosauravus moved to a more derived position. The elimination of a monophyletic Younginiformes is ironicallty more in keeping with the premise of transitional evolutionary grades (which also helps prune away some of those excessive ghost lineages). I am reminded here also of the difference in chelonian phylogenetic studies and cladograms between the earlier, ground-breaking work of Gaffney, and the more recent phylogenies of Joyce and others. In these and other instances, use of powerful computers as well as the benefit of more recent discoveries, and tend to key in as much data as possible, creating huge data matrixes and clades that cannot be easily identified by means of a few obvious synapomorphies. In these instance, it is hard to know which methodology should be preferred. In any case it seems that the relationships of these early diapsids are a lot less clear-cut then was previously thought.
One thing that everyone does agree on now is that the younginiforms preceded the great Lepidosaur - Archosaur split, and that there was a large evolutionary radiation during the Late Permian or, perhaps, Early Triassic (the latter dating makes more sense if we assume the Diapsids radiated to occupy the evolutionary niches vacated by the Synapsids and Parareptiles folliowing the end-Permian mass extinction) . Meanwhile there are other, more phylogenetically controversial taxa, such as the ichthyopterygia, sauropterygia, and many smaller groups, which may or may not be related to each other, and which may or may not belong outside or in one of the two main Diapsida groups. MAK111120
Sauria: LCA all living diapsids [C97a]
from the Middle Permian.
Neodiapsida:::: Ichthyopterygia + *: Archosauromorpha + Lepidosauromorpha.
Ant. process of squamosal narrow; squamosal largely restricted to top of skull; dorsal origin of temporal musculature; no tabular; sacral ribs lateral orientation; ribs mostly single-headed; fusion of caudal ribs, small proximal carpals & tarsals.
References: Callaway (1997a) [C97a] ATW990911.
Younginiformes: Acerosodontosaurus, Youngina, Hovasaurus, Tangasaurus. Medium-sized lizard-like, some forms aquatic. Possibly paraphyletic as ancestral to lepidosaurs.
Range: Late Permian to Early Triassic.
Neodiapsida::: (Ichthyopterygia + Sauria) + *.
Aquatic forms quite similar to terrestrial with laterally flattened tails. Single coracoid; tabular absent; quadrate not embayed; vertebrae with relatively tall neural spines; stapes large; sternum and vertebral transverse processes present; cleithrum present; olecranon process of ulna absent; primitive tarsus, pes; $ lateral centrale not in contact with 4th distal carpal; gastroliths.
Comments: Originally considered the most basal diapsids, this view was revised after discovery of Petrolacosaurus and other araeoscelids. Considered next lepidosauromorphs (Benton 1985; Evans 1988), moved to the base of Neodiapsida (Laurin 1991), now seem to be a praphyletic assemblage of basal forms (Bickelmann et al 2009). From the introduction:
The so-called "younginiform" reptiles (Benton 1985; Romer 1945) are known exclusively from the Upper Permian and Late Triassic of Madagascar and East Africa (Broom 1914; Carroll 1981; Currie 1980, 1981, 1982; Harris and Carroll 1977; Haughton 1924; Piveteau 1926). Originally thought to be the most basal diapsids, this view was modified after careful investigations of Petrolacosaurus and other araeoscelid reptiles (Reisz 1977, 1981; Reisz et al. 1984). Although "younginiform" reptiles were occasionally considered as lepidosauromorphs (Benton 1985; Evens 1988), Gaffney (1980), and later Laurin (1991) eventually placed them in their currently accepted position at the base of Neodiapsida. However, the in-group relationships of "Younginiformes," as well as their monophyletic status, are neither understood nor have they been tested in a modern phylogenetic framework. Currie suggested their monophyly in 1982, including Youngina capensis (Broom 1914), Hovasaurus boulei (Currie 1981), Tangasaurus mennelli (Currie 1982), Kenyasaurus mariakanensis (Harris and Carroll 1977), as well as Thadeosaurus colcanapi (Carroll 1981), but this interpretation was solely based on Hennigian argumentation, which was the only technique available at that time. The same methodology was also used in studies by Benton (1985) and Evens (1988) on diapsid "younginiform" phylogeny, in which both authors also included another taxon from the same deposits, Acerosodontosaurus piveteaui (Currie 1980). While Currie (1982) had surprisingly ignored this taxon in his phylogenetic study, both Benton (1985) and Evans (1988) placed it as sister taxon to all other "Younginiformes."
As defined here then the Younginiformes are simply a paraphyletic assemblage of broadly lizard-like (i.e. generalised reptilian) terrestrial and semi-aquatic basal Neodiapsid reptiles that flourished during the late Permian and early Triassic periods.
Links: Diapsids Phylogeny; Autapomorphies of diapsid clades. ATW010403. Palaeocritti, Wikipedia, Wikipedia Eosuchia, UCMP - Basal Diapsids (only a paragraph) MAK111118
Youngina: Youngina capensis Broom, 1914 (and other species?)
Synonyms: Acanthotoposaurus, Youngoides, Youngopsis
Late Permian (Tropidostoma-Dicynodon zones) of Southern Africa.
Links: Wikipedia
Tangasauridae: Acerosodontosaurus, Hovasaurus, Tangasaurus
Late Permian to Early Triassic (known from from the Sakamena Group of western Madagascar).
quadratojugal and jugal bones meet to form an arch in the skull, characteristic of many primitive diapsids, especially Younginiformes.. Smokeybjb 090221 (Wikipedia)
Comments: Relatively unaffected by the Permian-Triassic extinction event. (Ketchum & Barrett 2004) Benton 1985 divides the Tangasaurs into monphyletic Kenyasaurinae (terrestrial) and Tangasaurinae (aquatic). Kenyasaurinae is rejected by Bickelmann et al 2009, who however recovered a monophyletic Tangasauridae, excluding Kenyasaurus and Thadeosaurus but including Acerosodontosaurus. These were aquatic animals, with laterally compressed tails that allowed them to be able to swim in the freshwater lacustrine environments.
Links: Wikipedia

Late Permian of western Madagascar).
Comments: fully terrestrial, probably a strong runner. Included with Kenyasaurus as a monophyletic Kenyasaurinae by Benton 1985. Analysis by Bickelmann et al 2009 rejects the Kenyasaurinae and shows it to be a paraphyletic assemblage of basal Neodiapsids
Links: Wikipedia (incl. graphic)
checked ATW050512, revised MAK111120
Using this material. All material by ATW is public domain and may be freely used in any way (also any material jointly written by ATW and MAK). All material by MAK is licensed Creative Commons Attribution License Version 3.0, and may be freely used provided acknowedgement is given. All Wikipedia material is either Gnu Open Source or Creative Commons (see original Wikipedia page for details). Other graphics are copyright their respective owners