| Lepidosauromorpha | ||
| The Vertebrates | Lepidosauria |
| Vertebrates Home | Vertebrate | Vertebrate |
|
Abbreviated Dendrogram
○Diapsida
├─Archosauromorpha
└─○Lepidosauromorpha
├─Sauropterygia
└─○Lepidosauriformes
├─Kuehneosauridae
└─┬─Marmoretta
├─Megachirella
└─┬─Sophineta
└─○Lepidosauria
├─Sphenodontia
└─Squamata
|
Contents
Overview |
Although often overshadowed by their more charismatic big brothers, the archosaurs, the lepidosaurs are very much a success story in minature. An important element among micro- and meso-vertebrate faunas in the late Triassic and early Jurassic, when most lepidosaurs were rhynchocephalians, they underwent an extraordinary evolutionary radiation during the middle to late Jurassic with the appearance of the squamates. Today, lizard and snakes number over seven thousand species, second only to birds (Aves) as the most species-rich tetrapod group.
The Lepidosaurs emerged from an assortment of very lizard-like (in general appearance) lepidosauromorph/lepidosauriform taxa that flourished during the late Permian and early Triassic. During the early Triassic, these various basal forms gave rise to both the specialised gliding forms called kuehneosaurs, sauropterygians (probably), and the true Lepidosauria. All these reptiles doubt6less radiated into the ecological niches left by the end Permian extinction, which so devastated the mammal-like reptiles that until then had dominated terrestrial ecologies. This central group was mostly very small in size, focussing on small lizard niches that the archosauromorphs had not filled. Aong the stem of the main lepidosaur lineage (Crown Group Lepidosauria, to use cladistic terminology) were relict "living fossils" such as Marmoretta survived alongside lepidosaurs proper well into the Jurassic (Evans & Borsuk-Białynicka 2009 p.198).
The higher Lepidosaurs divided into two subgroups, the Sphenodontia (Sphenodon and its relatives) and the Squamata (lizards and snakes). The Sphenodontia, also called Rhynchocephalia abd Sphenodontida, were very common and successful from the late Triassic to the late Jurassic, but diminished progressively through the Cretaceous and Tertiary. The squamates in contrast, which are rare or unknown prior to the middle Jurassic, went from strength to strength as the Cretaceous and Cenozoic progressed. One is reminded of the pterosaurs being supplanted by the birds in the Cretaceous. The difference between the two groups, and perhaps the reason for the success of the lizard and snake line, can be explained in terms of their differences in cranial morphology, and especially the evolution of streptostyly (anteroposterior (forward and back movement of the quadrate bone) and skull and jaw mobility (cranial kinesis) in general.
Streptostyly is achieved in lepidosaurs through partial or complete loss of the strong lower temporal bar. This has occurred a number of times among different lines of diapsids, as has the development of a complete lower temporal bar. Biomechanical analysis shows that streptostyly reduces certain stresses acting on the skull (the quadratomandibular joint force) during biting, but at the cost of limiting maximum bite force and decreasing skull robusticity. Thus a mobile quadrate has the advantage of reducing the possibility of fracture, but an akinetic skull allows for a more powerful bite. The development of a lower temporal bar would provide additional support for an immobile quadrate which could become highly stressed during forceful biting. (Moazen et al 2009)
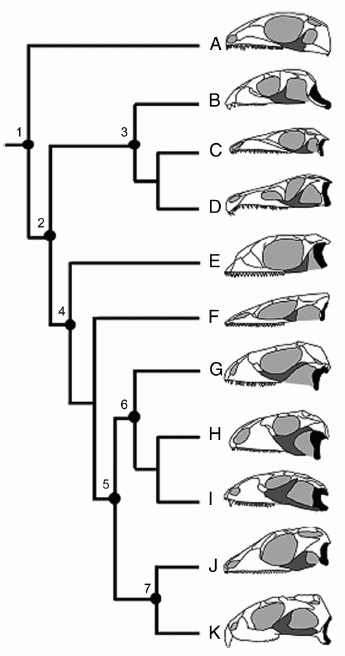
The following diagram shows some a range of variations and modifications of the lower temporal bar among diapsids and especially lepidosauriformes
|
|
A simplified phylogeny of diapsid reptiles showing variation in the development of the lower temporal bar. Black indicates quadrate; dark gray, jugal. (A) the basal diapsid Petrolacosaurus; (B) the early rhynchosaur Mesosuchus; (C) the archosauromorph Prolacerta; (D) the archosauriform Proterosuchus; (E) the lepidosauriform Kuehneosaurus ; (F) the stem lepidosaur Marmoretta; (G) the extant squamate Iguana, representing a more or less unspecialised type; (H) the large herbivorous Cretaceous scincomorph Polyglyphanodon, (I) the related late Cretaceous lizard Tianyusaurus, unique among squamates in having a rhynchocephalian-like akinetic quadrate; (J) the basal rhynchocephalian Gephyrosaurus; and (K) the advanced extant rhynchocephalian Sphenodon. Nodes: 1, Diapsida, primitively lower temporal bar, fixed quadrate, robust epipterygoid; 2, Sauria, no lower temporal bar, fixed quadrate and epipterygoid; 3, Archosauromorpha, no lower temporal bar in basal taxa, but developed in Rhynchosaurs and Archosaurs; 4, Lepidosauriformes and 5, Lepidosauria, no lower temporal bar in basal taxa quadrate, epipterygoid sutured to pterygoid, large squamosal; 6, Squamata, quadrate/pterygoid overlap reduced, ligamentous, squamosal reduced, epipterygoid with synovial pterygoid joint, development of lower temporal bar in Tianyusaurus only; and 7, Rhynchocephalia, no lower temporal bar in basal taxa, developed in some descendants, quadrate, pterygoid, and epipterygoid firmly sutured. Text (slightly modified) and diagram from Moazen et al 2009 p.8274 |
Rather than being Mesozoic relics or "living fossils" with the extant Tuatara representing a type unchanged since the late Triassic, the Sphenodontia constitute a diverse evolutionary lineage that included baqutic, herbivorous, and other forms (Benton, 1986). The two lepidosaur groups therefore each persued very different adaptive stratigies, one developing a strong, immobile, skull for powerful biting, the other a reduced and highly mobile skull. Sphenodontians developed a complete lower temporal bar perhaps several times, whereas ancestral squamates lacked a lower temporal bar and then underwent a reduction of other elements as well, and a reduction of the connections between them. In advanced Sphenodontia such as Sphenodon the quadrate, pterygoid, and epipterygoid extensively overlap and are firmly sutured together. In an advanced squamate such as Varanus, the contacts are substantially reduced, allowing greater mobility (Moazen et al 2009 p.8273), such as being able to move the upper jaw relative to the braincase. This ability is especially developed in snakes, and allows them to open their mouths extremely wide to swallow large prey.
Megachirella: Megachirella wachtleri Renesto & Posenato 2003
middle Triassic (Anisian) of Northern Italy
Comments: A stem Lepidosauriform, similar to Marmoretta. The species name honours Michael Wachtler, who discovered the specimen.
Links: - Megachirella detailed page and life illustration by Prof. Silvio Claudio Renesto. MAK101003
Reference: Renesto & Posenato 2003
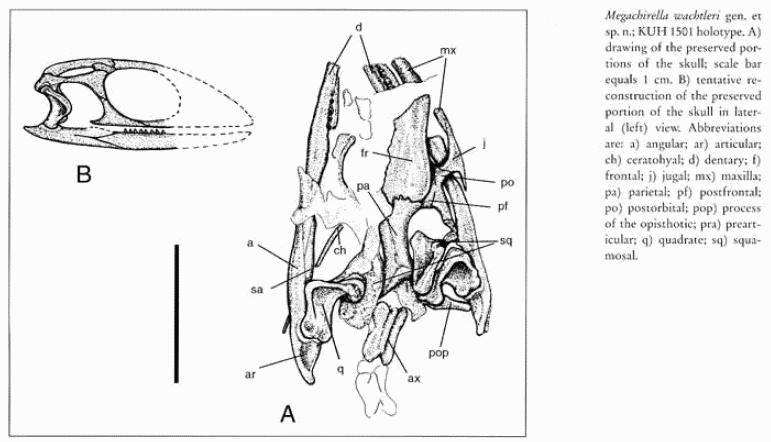 |
| Megachirella wachtleri, Anisian of Northern Italy. Diagram from Renesto & Posenato 2003 p.468 |
middle Jurassic (Bathonian) of England, related species at Guimarota, Portugal (Late Jurassic: Kimmeridgian) (Evans & Milner 1994)
Lepidosauriformes: (Sophineta + Lepidosauria) + *.
Comments: Relict Jurassic genus, shares primitive features in common with kuehneosaurs. Possibly aquatic.
Reference: Evans, 1991
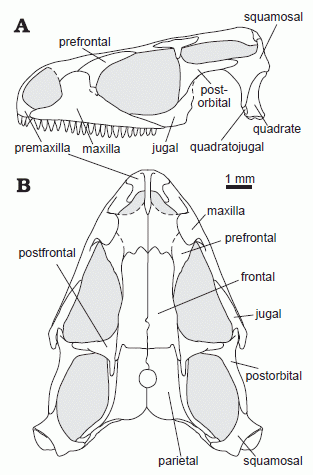 |
| Sophineta cracoviensis, Olenekian of Poland. Skull reconstruction in lateral (A) and dorsal (B) views. Diagram from Evans & Borsuk-Białynicka 2009 p.182 |
Sophineta: Sophineta cracoviensis Evans & Borsuk-Białynicka 2009
Early Triassic (earliest Late Olenekian) of Poland
Lepidosauriformes: Marmoretta + (Lepidosauria + * ).
Characters: small (skull ~ 1cm, body with-out tail ~3 cm), relatively short preorbital region, tall facial process of the maxilla, reduced lacrimal, and pleurodonty, all derived features that resemble early crown-group lepidosaurs rather then stem-taxa.
Comments: sister taxon of Lepidosauria, shares features in common with the basal rynchocephalian Gephyrosaurus ( Evans & Borsuk-Białynicka 2009)
Range: from the Late Triassic
Phylogeny: Lepidosauriformes : Kuehneosauridae + (Marmoretta + (Sophineta + * : Rhynchocephalia + Squamata).
Characters: Determinate growth with epiphyses on terminal ends of long bones; post-parietal and tabular absent; fused astragalus and calcaneum; kidney sexual segment; thin stapes. Thyroid fenestration. Significance: hearing (lost in sphenodontids, strong foot action & ability to jump?
checked ATW031006, last revised MAK101030