 The Dichobunids: Variations on a theme of Diacodexis
The Dichobunids: Variations on a theme of Diacodexis| Cetartiodactyla | ||
| The Vertebrates | Artiodactyla |
| Vertebrates Home | Vertebrate | Vertebrate |
|
Abbreviated Dendrogram
Laurasiatheria
├─Ferae
└─┬─Perissodactyla
│
└─Cetartiodactyla
├─Cetacea
│ ├─Pakicetidae
│ └─Autoceta
└─Artiodactyla
├─Dichobunidae
└─┬─Suina
│ ├─Raoellidae
│ └─┬─Hippopotamidae
│ └─Suoidea
└─Selenodontia
├─Camelidae
└─Ruminantia
├─Giraffoidea
└─┬─Bovoidea
└─Cervoidea
|
Contents
Overview |
In the long run, the demise of the ornithischian dinosaurs principally benefited two groups: the rodents and the ungulates. Of the ungulates, the dominant group is now the artiodactyls. This hasn't always been so. The perissodactyls were once much more numerous. But the artiodactyls were able to adapt and thrive in the harsh climatic conditions which have prevailed since the Late Miocene. The principle advantage of the artiodactyls is sheer efficiency at turning low quality fodder into flesh. Other mammals are faster, stronger, smarter, more agile, or more adaptable. The artiodactyls, in fact, tend to respond to environmental change mostly by dying in large numbers. However, after the fire, flood, ice age, drought, contamination, or volcano has run its course, the region will be first recolonized by grasses and adventitious weeds. If any artiodactyls are left in the region at all -- they range far and quickly -- they will return before any others, as the large mammals best able to make a living off this marginal ground cover. Their herds will rebuild faster than potential competitors can re-establish their more demanding life-styles. As a result, any catastrophe which does not completely erase the artiodactyls will only make them stronger. ATW040723.
The Dichobunids: Variations on a theme of DiacodexisThe dominance of the artiodactyls would have been hard to predict in the Early Eocene when the main ungulate clades diverged. The earliest known artiodactyls are referred to as the Dichobunidae. This is probably a paraphyletic group of basal artiodactyls. Most are variations on Diacodexis. These rabbit-sized animals were among the most cursorial animals of their time, and may have been capable of rabbit-like leaping. If we may compare them to any modern artiodactyl, we might think of them as tiny, hyperactive goats. Their artiodactyl specializations were almost all in the limbs -- in particular, the characteristic artiodactyl "double-pulley" astragalus, which has a functional trochlea at both ends. However, even this may be misleading. The most basal species of Diacodexis comes from South Asia. This species lacks even the cursorial adaptations of the better-known North American form. So, either Diacodexis is the actual parent of all other artiodactyls, or its adaptations for running were evolved convergently. Currently, the belief is that this is a case of convergence.
The artiodactyls then split into the Suina and the Selenodontia. The suines are pigs, hippos and their close relatives -- maybe. Molecular phylogenists, for once allied with some very able morphologists, have suggested that the whales may have split off from within the suines, rather than earlier. If so, Artiodactyla (currently deer > dolphins) will probably go back to being a crown group (deer + pigs). However that may be, it might be more useful to think of this basal split among artiodactyls as selenodonts vs. everyone else. The selenodonts have an impressive number of specializations, and will be taken up later. The suines are consummate generalists. ATW040723.
 Suina: Anything Goes
Suina: Anything GoesCollectively and individually, the suines are extraordinary generalists. They will eat almost anything and live almost anywhere. Since they tend to be almost hairless, they don't do well in really cold climates, but their wonderfully insulated bodies are quite comfortable virtually anywhere else, either on land or water. In addition, some of them are (as George Orwell noted), the intellectual leaders of the artiodactyls, although they can also be destructive and aggressive. They tend to be gregarious, but their social behavior is flexible. The parallels between pigs, in particular, and humans are striking. This similarity was not lost on our own ancestors, with important cultural echoes in many human societies. Suines retain the strikingly triangular skull shape of Diacodexis as well as the relatively unspecialized limbs, bunodont molars, canines, and other very ancient features. The dentition tends to develop strong sexual dimorphism, with large lower canine tusks on males.
The broad environmental tolerance of suines comes, however, at a price. That price is the very absence of the selenodont specializations, the most obvious of which is foregut fermentation. Suines are not ruminants. They are capable of eating their vegetables only because of an enlarged cecum,a pocket between the small and large intestine in which they house bacteria capable of breaking down cellulose. This procedure is considerably less efficient -- albeit also less disgusting -- than the foregut fermentation of ruminants (see figure under Artiodactyla) [PS02]. The difference in efficiency may be more than offset by other characteristics. For example, ruminants may take many days to fully digest their food, and the heat generated by fermentation on this scale can be a serious physiological liability [PS02]. As a result, pigs, for example, are notably more efficient than cows at turning greens into meat when measured as kg meat/kg greens/month under primitive agricultural conditions [W04]. However, a cow can eat enormous quantities and digest on the move. A pig has to eat almost every day. This has made an enormous difference to human history. Cattle can be domesticated by nomads in marginal environments. Pigs cannot. We will take up more of this interesting topic, and its implications, in due course. ATW040723.
There's a theory in circulation that artiodactyls derived from Indian stock -- possibly picked up in Africa, as the subcontinent passed East Africa on its way from Gondwana to Asia. So, for example, the first and most primitive form of Diacodexis appears suddenly in Pakistan in the Early Eocene, just about the time India docked with the mainland. Likewise, the Raoellidae appear to the east of India not much later. Either they represent an early radiation of Indian dichobunid-like basal stock or perhaps they had already diverged in India. Unfortunately, most of India was either under water or reworked by volcanic basalts during the latest Mesozoic and Paleocene, while most of the north coast is now buried far beneath the Himalayas -- so its hard to tell.
One peculiar feature of raoellids is molar wear pattern including near-vertical gouges. Such marks are also known from the pakicetids -- and almost nowhere else. The same pattern is found in Georgiacetus, a Middle Eocene whale from North America. Thus, it is somewhat difficult to dismiss this characteristic -- as many do -- as something unknown but unique to Early Eocene India. Raoellids may be poorly known, but we have no reason to believe that they were aquatic to the same degree as whales. ATW040724.
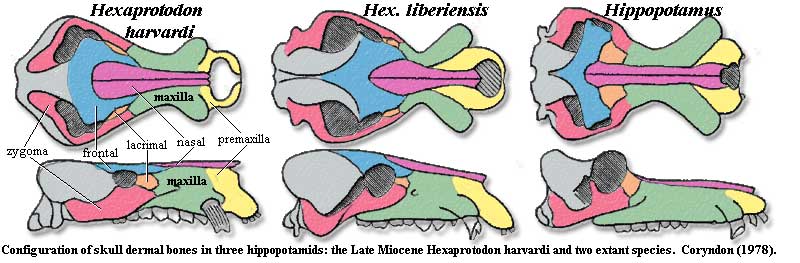
Some of the most basal suines may be the extant Hippopotamidae. While we have placed them as primitive suines, there are several other theories. The hippopotamids seem to have been rather marginal creatures during most of suine evolution, and became common only in the latest Miocene and Early Pliocene [C88]. They show a number of typical adaptations for aquatic life, such as dorsal orbits and nares and somewhat shortened limbs. Unfortunately, this may give a misleading phylogenetic impression.
Traditionally, they were regarded as an offshoot of the anthracotheres because some of the anthracotheres probably had a very similar lifestyle. Another theory, perhaps the most widely held today, is that hippos are peccaries Tayassuidae), and thus deep within the Suoidea [PS02] [P95]. But, if whales are close to hippos, and hippos are close to peccaries, then whales are suoids -- which is as close to pure nonsense as we are likely to see out of sensible paleontologists. Something has to give, and -- since the fossil record of hippos is notoriously poor -- the weakest link is the hippo-peccary connection. ATW040724.
 Artiodactyla: Diacodexis, pigs, camels, deer, cattle.
Artiodactyla: Diacodexis, pigs, camels, deer, cattle.
From the Early Eocene [C88].
Cetartiodactyla : Cetacea + * : Dichobunidae + (Suina + Selenodontia).
Most successful ungulate group; long preorbital skull formed by elongated frontals; frontals commonly bear horns; postorbital bar present; premolars not fully molariform and frequently specialized; canines retained and often large [C88]; P1 double-rooted [T+01]; primitively, p3 not reduced [C88]; molars bunodont or selenodont; primitively, all M without hypocone (only in most basal forms) [C88]; M2 metacone and paracone similar height [T+01]; $ M2 trigon basin large [T+01]; $ M2 paraconule present [T+01]; M2 metaconule present and similar in size to paraconule [T+01]; $ m2 trigonid and talonid roughly equal size [T+01]; $ m2 hypoconulid & entoconid present [T+01]; jaw condyle not far above tooth row [C88]; glenoid fossa without preglenoid tubercle [T+01]; ectotympanic forms part of external auditory meatus [T+01]; 4 - chambered stomach as shown in figure, with chambers I-III derived from esophagus; plant material initially held in rumen and gradually digested (slower digestion favors medium-large forms with lower metabolism very large forms have low enough metabolism to rely on intestinal fermentation); clavicle usually absent; $ acromion process of scapula does not overhang glenoid [T+01]; distal ulna and fibula may fuse with radius or tibia; "double pulley" arrangement of astragalus, with pulley shaped articulations in same plane with tibia and tarsals (or carpals) allowing no lateral rotation of foot [C88]; springing ligaments; $ distal phalanges narrow in dorsal view [T+01]; paraxonic foot, with plane of symmetry between digits III & IV, which are the primary weight-bearing elements; digits II and V usually reduced or absent, digit I generally absent; metapodials III & IV frequently fused as cannon bone; very often with hooves.
Links: Introduction to the Artiodactyla; Animal Diversity Web: Order Artiodactyla; Artiodactyla; Artiodactyls: AllAboutMammals.com; Artiodactyl Identification; Erfurt's fourth Page; Vertebrate Paleontology Photo Gallery; Artiodactyla; Artiodactyla; Order ARTIODACTYLA; Order Artiodactyla; Mammalia/UNGULATA/artiodactyla.html">Mammalia, Ungulata, Artiodactyla - Paarhufer; artiodactyla page; Comparative Mammalian Brain Collections: ARTIODACTYLA; Introduction to Artiodactyla; Bibliography of Ungulates.; Artiodactyla; ORDO ARTIODACTYLA OWEN, 1848 (Russian & English: karyotype data & references).
References: Carroll 1988) [C88], Thewissen et al. (2001) [T+01]. ATW040814.
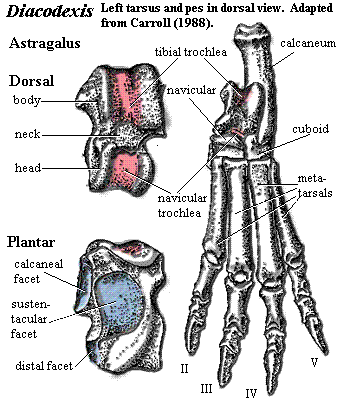
 Dichobunidae: Bunophorus, Diacodexis, Dichobune, Hyperdichobune.
Dichobunidae: Bunophorus, Diacodexis, Dichobune, Hyperdichobune.
Range: Early Eocene to Late ?Oligocene of India (Diacodexis), Europe, Asia & North America. [C88]
Phylogeny: Artiodactyla : (Suina + Selenodonta) + *.
Characters: simple bunodont cusps [C88] [PS02]; complete set of teeth [C88]; upper premolars simple (i.e. not molarized) [SL00]; P3 &
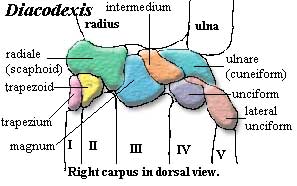
4 mesiodistally elongate [SL00]; p4 with moderately strong paracristid [C88]; upper molars retain triangular cusp pattern [C88]; lower molars with talonid lower than trigonid [SL00]; hypoconulid absent [SL00]; m1 & m2 with talonid wider than trigonid [C88]; clavicle present primitive) [C88]; limbs elongate; carpus with additional, 5th element in distal row of carpal bones ("lateral unciform") (primitive); metapodials elongate, not fused, but closely integrated [C88]; five complete digits in manus; hind limbs elongate [C88]; femur retains 3rd trochanter [C88]; tibia longer than femur [C88]; fibula splint-like [C88]; double pulleyed astragalus [C88]; astragalus, tibial trochlea shallow (primitive); sustentaculum short primitive); astragalus, cuboid = distal) facet narrow (~30% of 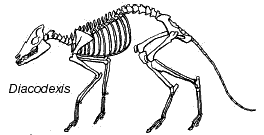 astragalus distal width?); navicular not fused with cuboid (primitive) [C88]; four digits in pes; digitigrade.
astragalus distal width?); navicular not fused with cuboid (primitive) [C88]; four digits in pes; digitigrade.
Image: Diacodexis carpus after [SL00].
Links: Dichobunidae Mikko's Phylogeny); Erfurts's Abstracts; Erfurt's fourth Page; New Morphological Evidence for the Phylogeny of Artiodactyla ....
References: Carroll (1988) [C88], Prothero & Schoch 2002) [PS02], Sudre & Lecomte (2000) [SL00]. ATW040814.
 Suina: = Suiformes. pigs > camels. pigs & hippos
Suina: = Suiformes. pigs > camels. pigs & hippos
Range: from the Early Miocene
Phylogeny: Artiodactyla :: Selenodontia + * : Raoellidae + (Hippopotamidae + (Anthracotheroidea + (Entelodontoidea + Suoidea))).
Characters: unique skull shape; squamosal extends to the exoccipital, covering the mastoid in lateral view [C88] [1]; canines with marked sexual dimorphism; bunodont molars; unspecialized premolars; short diastema; retain fibula & ulna; 4 digits; stomach simple
Image: Babyrousa from the Ungulate Collection of the University of Edinburgh Natural History Museum.
Notes: [1] may apply to a slightly less inclusive clade.
Links: suina page very good short introduction); Suina Mikko); Chapter 1- Action Plan systematics of living forms); artiodactyles French).
References: Carroll (1988) [C88], Prothero & Schoh (2002) [PS02], White (2004) [W04]. ATW040723.
Raoellidae: previously grouped with Helohyidae. Haqueina, Indohyus, Khirtharia, Kunmunella, Raoella
Range: Early Eocene, but mostly Middle Eocene of South and Southeast Asia. May have been present in Late Eocene.
Phylogeny: Suina : (Hippopotamidae + (Anthracotheroidea + (Entelodontoidea + Suoidea )) + *.
Characters: dental wear patterns as near-vertical gouges (as in Pakicetids?) [T+00].
Links: Eocene mammal faunas from northern Indo-Pakistan; PaleoNET | Библиотека; .
References: Thewissen et al. (2001) [T+01]. ATW040721
 Hippopotamidae: Hexaprotodon (= Choeropsis), Hippopotamus, Kenyapotamus
Hippopotamidae: Hexaprotodon (= Choeropsis), Hippopotamus, Kenyapotamus
Range: from the ?Early Miocene of Europe, Asia and Africa. Become relatively common only in the latest Miocene and Early Pliocene. [C88]
Phylogeny: Suina :: (Anthracotheroidea + (Entelodontoidea + Suoidea)) + *.
Characters: relatively large (to 3600 kg); skull very large and elongated; nares dorsal; well-developed sagittal crest; orbits dorsal; incomplete postorbital bar; braincase small; jaw broad & square; dental formula primitively complete [C78]; extant species 2-3/1-3, 1/1, 4/4, 3/3 [1]; incisors & canines ever-growing; lower incisors procumbent & longer than uppers; lower canines very long & project above upper tooth row; molars bunodont, but tend to wear down in somewhat loph-like patterns; first molars erupt early and are worn almost completely before eruption of 2nd & 3rd [C78]; M3 with tall distal cingulum [C78]; m3 with distinct talonid [C78]; bodies rounded; tail short; legs short & thick; limb bones not fused; four functional digits on all limbs; lateral digits not reduced; digitigrade; stomach with three chambers (but not ruminating); skin thick & mostly) hairless; amphibious and gregarious.
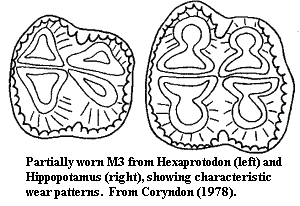 Notes: [1] There seems to be an unreasonable amount of disagreement on this point. Coryndon [C78] gives the formulla of extant species as 2/1-2, 1/1, 3-4/3-4, 3/3 (with first premolars present only as milk teeth), the lower numbers always referring to Hexaprotodon.
Notes: [1] There seems to be an unreasonable amount of disagreement on this point. Coryndon [C78] gives the formulla of extant species as 2/1-2, 1/1, 3-4/3-4, 3/3 (with first premolars present only as milk teeth), the lower numbers always referring to Hexaprotodon.
Links: Hippopotamidae Mikko's Phylogeny); Animal Diversity Web: Hippopotamidae: Classification excellent site for extant species); Family Hippopotamidae the usual excellent skull photos from the Tokyo Univ. School of Medicine); ARTIODACTYLA; HIPPOPOTAMIDAE; HIPPOPOTAMUS- Hippopotamus on-line version of Walker's Mammals); ECOLOGY PHOTOGRAPHIC Hippopotamidae photos of hippos under water); artiodactyls; A ... images); Hippopotamus; Mammalia/UNGULATA/HIPPOPOTAMIDAE/hippopotamidae_01.html">Flußpferde - Hippopotamidae German); Hippopotamuses! (GeoZoo); Hipopótamos.
References: Carroll (1988) [C88], Coryndon (1978) [C78], Prothero & Schoh (2002) [PS02], Petronio 1995) [P95]. ATW040721.
checked ATW040705