|
|
Bones: The Gill Arches |
| The Vertebrates |
The Epibranchials |
The Gill Arches: The Epibranchials
 Summary Summary
The epibranchials are the main upper elements of the gill arches. Fish with jaws usually have five functional pairs of gill arches. The last arch is sometimes simplified, so the typical fish has either 4 or 5 pairs of epibranchials. The lower ends of the epibranchials are attached to the ceratobranchials. The upper ends attach to pharyngobranchials or some related bone. The epibranchials may also support comb-like gill rakers, tooth plates for pharyngeal (throat) teeth, or various flanges for the attachment of muscles. The most important soft tissue attachments are the gills themselves, which project outward from the epibranchials, opposite the rakers and/or pharyngeal teeth. The epibranchials also have a variety of soft tissue attachments to the wall of the throat, gill muscles, and any interbranchial septa (tissues separating the gill chambers).
The epibranchials have an interesting long-term evolutionary relationship with the ceratobranchials. Generally, the epibranchials follow "Williston's Law." That is, the number of epibranchials decreases and their degree of individual specialization increases. However, this seems to occur by shifting the core respiratory function onto the ceratobranchials, which change very little.
The epibranchials can perhaps be traced back into the jawless, osteostracan fishes. In sharks and actinopterygians we see some very similar trends, including (a) reduction in size (b) cross-linking or specialization to support terminally cross-linked pharyngobranchials, and (c) loss of 1-2 posterior epibranchials. In the teleosts, the epibranchials also develop a series of processes and flanges for the attachment of branchial muscles. |
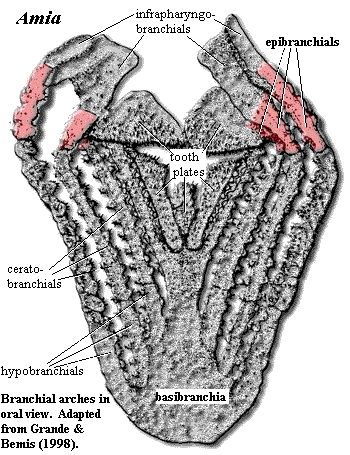 The Standard Condition
The Standard Condition
To the left is a nonstandard view of the Standard Condition, as exemplified by our usual standard in matters branchial, Amia calva. Here we are staring down the throat of Amia, a perspective normally obtained only by unfortunate prey items in their final moments before maceration on the pharyngeal tooth plates lining the throat. Petty distractions, such as dermal bones, jaws, and the hyoid arch, have been removed. Amia, as has been observed elsewhere, is well-endowed with teeth -- on the jaws, on the palate, and abundantly on the gill arches.
In Amia as in many other fishes, there are five branchial arches, but only four epibranchials. The first two epibranchials support infrapharyngobranchials which are fused dorsally on each side. Epibranchials 3 & 4 support the massive upper tooth plates. In Amia, as in many actinopterygian fishes, the ceratobranchials do much of the mundane work and heavy lifting. The epibranchials are short and appear more concerned with higher matters, such as the support and orientation of a variety of other dorsal elements, the infrapharyngobranchials and additional tooth plates.
This is an interesting example and counter-example of Williston's Law at work. Williston's "Law" is that, in any given lineage, the number of serially homologous elements tends to decrease while the individual elements tend to become more different and specialized. Some of the best examples are the cranial bones and the teeth of mammals. Of course, there are no "laws" in paleontology equivalent to those in the harder sciences, but it is a common pattern. Here, the number of epibranchials has been reduced, and their functions have diverged somewhat from each other, in accordance with the "law." However, the epibranchials' original core function of supporting the gills is not one that can easily be abandoned. Instead, this function has been shifted progressively shifted to the ceratobranchials which have retained the ancestral number (5) and remain quite uniform in morphology.
One suspects that this is one of those chaotic processes in which a small random event has set in motion an irreversible cascade of consequences. For, as soon as any epibranchial is slightly adapted to functions other than gill support, two things happen. First, selective pressure increases on the corresponding ceratobranchial not to diverge, but to take up the respiratory slack. Second, the epibranchial's divergence must result from some interference in the regime of genetic regulation which originally caused it to develop as a serial structure in the first place. The breaking of this genetic assembly line for one serial element necessarily means that it is more easily possible to divert other serial elements, since all are created from the same package of genes. Thus, each change increases the tendency of epibranchials to diverge, and the tendency of ceratobranchials not to diverge.
That second point is rather abstract. Let's try a molecular biology example. Suppose a very slightly aberrant regulatory element in a structural gene related to the epibranchials becomes slightly sensitive to some universal regulator -- like one of the bone morphogenetic proteins (call it BMP-x). Subsequently, an ancestor develops with slightly elevated levels of BMP-x during development, near epibranchial 2. The result is a slightly misshapen bone which, perhaps, has some slight advantage in allowing the pharyngobranchial elements to converge and fuse (as in Amia). If that animal were to live long and prosper, the structural gene in its descendants will tend to evolve a stronger and more consistent quantitative response to BMP-x. But the same gene is used in all of the serially homologous epibranchials. Thus, every member of the entire series becomes sensitive to relative levels of BMP-x in its particular developmental environment. This sensitivity, in turn, renders all the epibranchials subject to regulation by BMP-x antagonists and enhancers so that divergence becomes increasingly likely. In the process, the epibranchials will likely become less optimally adapted to gill support, which puts selective pressure on the ceratobranchials to focus all the more strongly on their core function. The result of the original tiny mutation, is a cascade of events in two directions, with the epibranchials becoming ever more individualized and the ceratobranchials becoming increasingly dour and conformist.
Phylogeny
Jawless Fishes: The history of the epibranchials presumably begins wherever vertebrates first developed a jointed, internal branchial arch rather than an unjointed, external branchial basket. In thelodonts, perhaps the denticle patches with fused bases found in the pharyngeal area near the presumed gills of loganiids Marss & Ritchie (1998); Van der Brugghen & Janvier (1993)) are the first possible indication. If so, this is particularly interesting because the reconstructions of the furcacaudiform thelodonts suggest branchial bars of the external variety. Wilson & Caldwell (1993); Wilson & Caldwell (1998). Certainly osteostracans are supposed to have had internal gill arches, either as part of, or supported by, the cranial cartilage. Janvier 1996).
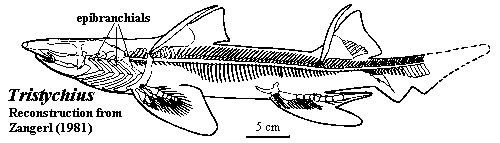 Placodermi and Chondrichthyes: Very little is known about the branchial arches in early gnathostomes. Placoderms, in particular seem to have had branchial elements which were almost entirely cartilaginous. Janvier (1996). As for sharks, "[t]he branchial arches are so poorly known in Paleozoic forms that little need be said about their specific morphology." Zangerl 1981: 26). To the extent that these structures are known, the gill arches of early Chondrichthyes seem to have had a classically symmetrical, 'V' shape with apex of the 'V' posteriorly directed. The epibranchials were then relatively long, straight elements. The word relatively should be stressed, since the entire branchial apparatus tended to be quite small and still compressed under the braincase, as in living Holocephali. This morphology remains consistent into the elasmobranchs and Neoselachii, but the entire apparatus becomes much larger and is located much further back, largely behind the neurocranium. Id. (The image shows a somewhat intermediate form, Tristychius, a Carboniferous (Viséan) elasmobranch. Here, the branchial arches are large, but are still closely related to the braincase.)
Placodermi and Chondrichthyes: Very little is known about the branchial arches in early gnathostomes. Placoderms, in particular seem to have had branchial elements which were almost entirely cartilaginous. Janvier (1996). As for sharks, "[t]he branchial arches are so poorly known in Paleozoic forms that little need be said about their specific morphology." Zangerl 1981: 26). To the extent that these structures are known, the gill arches of early Chondrichthyes seem to have had a classically symmetrical, 'V' shape with apex of the 'V' posteriorly directed. The epibranchials were then relatively long, straight elements. The word relatively should be stressed, since the entire branchial apparatus tended to be quite small and still compressed under the braincase, as in living Holocephali. This morphology remains consistent into the elasmobranchs and Neoselachii, but the entire apparatus becomes much larger and is located much further back, largely behind the neurocranium. Id. (The image shows a somewhat intermediate form, Tristychius, a Carboniferous (Viséan) elasmobranch. Here, the branchial arches are large, but are still closely related to the braincase.)
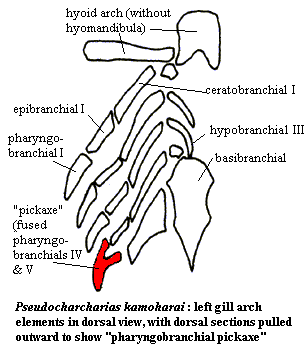 Even in Tristychius, the epibranchial has lost the secondary function it performed in the Osteostraci, that of fixing the gills to the cranial cartilage. That function has been taken over by specialized pharyngobranchials. By the level of the extant lamniform sharks, we see some of the same specializations found, convergently, in the Standard Condition: epibranchials of reduced size, supporting terminally fused pharyngobranchials, and loss of 1-2 posterior epibranchials. Curiously, this same condition is found in the rays and skates (Hypnosqualea), despite a very different body form. Compagno 1999a). These may be primitive traits for Neoselachii; but their persistence, and the degree of convergence with Amia, are initially startling.
Even in Tristychius, the epibranchial has lost the secondary function it performed in the Osteostraci, that of fixing the gills to the cranial cartilage. That function has been taken over by specialized pharyngobranchials. By the level of the extant lamniform sharks, we see some of the same specializations found, convergently, in the Standard Condition: epibranchials of reduced size, supporting terminally fused pharyngobranchials, and loss of 1-2 posterior epibranchials. Curiously, this same condition is found in the rays and skates (Hypnosqualea), despite a very different body form. Compagno 1999a). These may be primitive traits for Neoselachii; but their persistence, and the degree of convergence with Amia, are initially startling.
Acanthodii and Actinopterygii: An image of the acanthodian gill arches can be seen in the discussion of the Hypohyal. The basic morphology of the epibranchial is almost identical to Tristychius. Note, however, the presence of tiny ossifications along the shaft. These are gill rakers. Gill rakers are found in both chondrichthyans and in Osteichthyes, as well as acanthodians. Although no one seems willing to swear that rakers are homologous in all three groups, it does seem likely [1]. Perhaps gill rakers are also the forerunners of the pharyngeal teeth in the Standard Condition, although many advanced teleosts (e.g., labroids) have both, and the two populations of tooth-like bones seem to be distinct. Barlow (2000).
We are straying somewhat from the epibranchial itself, but it is all for a good cause. The primary practical importance of the epibranchial in fish work is as the bearer of gill rakers and various dorsal structures. The fine structure of fish phylogeny, particularly among the teleosts, is built on such distinctions. Even to experts, one anchovy looks very much like another. Thus, distinctions are diagnosed by the size and number of gill rakers and the presence or absence of rakers on the posterior face of the third epibranchials.
Neopterygii and Teleostei: The tendency to cross-link certain arches dorsally continues. Typically this occurs through fused pharyngobranchials, as in Amia. However, the epibranchials also become directly involved. For example, an interarcual cartilage frequently connects the first epibranchials with the second infrapharyngobranchials. In cyclosquamates, an elongated (uncinate) process of the second epibranchial contacts the third pharyngobranchial. Another teleost refinement is the levator process on the epibranchials, presumably for the levator brachialis muscles, as well as the uncinate process mentioned above. The presence, absence, size and position of these elements can be of considerable significance. See, generally, Esociformes, Wilson & Veilleux (1982), Johnson & Patterson (1997).
What all this might be telling us is that the gill arches are being recruited directly to assist in the active pumping of water across the surface of the gills, maceration by the pharyngeal teeth, and similar matters. Generally, the burden of adapting to these functions is falling on the dorsal elements for the reasons discussed above in connection with the Standard Condition [2]. The epibranchials thus (a) become shorter, increasing their mechanical advantage as levers to move the ceratobranchials, (b) cross-link, to improve resistance to muscular stresses, and (c) develop flanges and processes which facilitate muscle attachment and fine control of the more dorsal elements, such as the tooth plates. This reaches something of a logical extreme in cichlids, with their fully developed pharyngeal jaws. Barlow 2000).
Sarcopterygii & Tetrapoda: Even applying our customarily debased threshold of expertise, we lack sufficient knowledge of the sarcopterygian branchial apparatus to say much of anything meaningful. Since sarcopterygians developed neither pharyngeal teeth nor the advanced suction feeding methods of the teleosts, the general structure of the gills seems to have remained primitive. If one's basic feeding strategy is to get the largest possible mouth around the biggest possible food item and move it into the stomach as quickly as possible, then all anatomical subtlety is wasted. Our ancestors' lack of branchial refinement merely reflects their absence of manners or sophistication in matters of feeding.
As most sarcopterygian groups were moving (or being pushed) toward shallow, deoxygenated waters or even onto land, the utility of gills for ventilation decreased. Acanthostega, although fully aquatic, had only three, relatively modest, pairs of functional gills. It may have used its lungs to obtain most of its oxygen, with the gills specialized to shed excess carbon dioxide. Clack 2002). Perhaps for this reason, some basic gill structures survived for a very long time, particularly in the temnospondyl lineages. Id.
Crown Tetrapoda*: In most tetrapods*, the epibranchials lose their separate identity and are merged into structures in which their homology cannot be accurately determined. The first epibranchial is incorporated into the posterior horn of the hyoid. It sometimes emerges from this obscure retirement post to form a dramatically enlarged portion of the tongue skeleton in tetrapods* with extensible tongues (e.g., birds and salamanders). The other epibranchials are lost or merged tracelessly into the thyroid or tracheal rings. ATW030830.
[1] We know of no evidence for gill rakers in placoderms, but so little is known of placoderm branchial arches that it is hard to exclude the possibility. I have always considered it particularly weird that Bothriolepisand related antiarchs have structures on the medial face of the pectoral "limb" which look for all the world like gill rakers. This is one more indication of a deep, if recondite, relationship between the pectoral fins and the gills.
[2] This tendency is also related to the trend for the hypobranchials to transform from stick-like extensions of the ceratobranchials into rounded bearings on which the ceratobranchials can move relative to the basibranchium. See, the Hypohyal.
checked ATW030122