Placodermi: Antiarcha
Taxa on This Page
- Antiarcha X
- Asterolepidoidei X
- Bothriolepidoidei X
- Chuchinolepidae X
- Euantiarcha X
- Microbrachiidae X
- Minicrania X
- Sinolepidae X
- Yunnanolepidae X
- Yunnanolepidoidei X
Descriptions
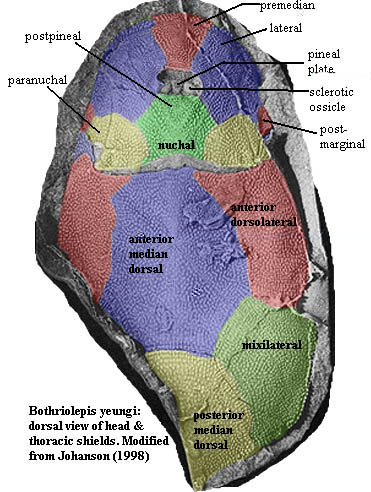
 |
Bothriolepis yeungae modified from Johanson 1998). To suit my own ill-informed prejudices on the matter, the tail has been reduced in size, and the body and head have been adjusted to suggest a more vertical orientation of the lateral plates. The changes made here are intended to conform the dorsal view more closely to Stensiö's reconstruction of the overall body form. |
Antiarcha:
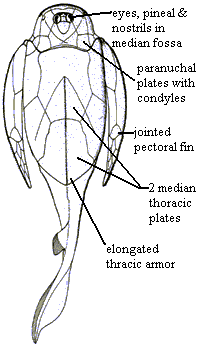
Head short; no superognathals (Carroll (1988) says 1 set; Janvier 1996) says this is the suborbital plate. In any case, 1 set of plates bites against infraognathals); eyes, pineal & nostrils housed in a common median dorsal fossa (or fenestra); orbital fossa primitively far anterior; fossa spiracular opening; endolymphatic ducts in pentagonal nuchal plate (median cephalic plate); nuchal plate with 2 outlets for endolymphatic ducts (open to environment as in chondrichthyans); $ unique skull roof plate pattern; head articulation involves cephalic condyles on paranuchal plates & fossae on thoracic dorsolateral plates (reverse of most other placoderms); elongated trunk armor; $ 2 median dorsal thoracic plates; pectoral appendages encased in bone; in derived forms, appendage is jointed & develops complex joint w axial exoskeleton; $ pectoral fin covered in dermal plates; $ pectoral fenestra contained in 1 plate; small, unplated pelvic fins.
Links: Faktasider Norwegian); Paleontology and Geology Glossary; Bothriolepis sp. (see also other links in the Paleontological Museum); Phylogeny of Antiarch placoderms (pdf/poster) - Sarah Kearsley.
Note: Remained heavily armored, benthonic; restricted to marginal marine or deltaic areas and FW; appendage used to cover fish with sand? 010626.
Discussion:
I. Canowindra: the Age of Fishes and the Road to Gooloogong
Canowindra is a small town of 1200 in the back country of New South Wales, in southeastern Australia, a few hundred km west of Sydney. The town was founded at a ford on the Belubula River. During the nineteenth century, it served as a road head for farmers and miners operating in the area. However (according to the tourist literature at any rate) it was much more famous as an outlaw town, and served as the Australian equivalent of the American Dodge City. More recently, Canowindra has billed itself the "Ballooning Capital of Australia," referring not to its size or population, but to its major involvement with hot air ballooning.
Perhaps it is the legacy of Ben Hall, a bush ranger who essentially kidnapped the entire town in 1863, or just something in the water; but the town of Canowindra has had a history of self-promotion which would do honor to a city a hundred times its size. It seems to have almost as many promotional web sites as adult citizens. The restoration of the downtown buildings, maintenance of tourist facilities and the hot air balloon festival represent a collective investment and organizational effort equivalent to the city of Birmingham (UK or Alabama -- either or both) getting together to put a man on the moon.
What does this have to do with paleontology? In 1956, the government of NSW decided to reward a contractor by putting in a paved road between Canowindra and the tiny suburb of Gooloogong (pop. 250), a town so small it isn't on most maps, and is spelled in an awkward variety of ways when it does appear. The road still isn't finished (according to the same maps). However, an errant bulldozer on the project turned up a peculiar slab of rock which was literally full of placoderm fossils. By a weird twist of fate, the find was recognized as important and immediately shipped off to Sydney. Much later, during the 1980's and early 90's, placoderm guru Dr. Alex Ritchie tried to find and expand the site, without much luck. Then, in 1992, he was invited to give a little talk on the subject to the Canowindra Rotary Club.
 In most small towns, this Rotary Club talk would have would have been the non-event of the season. But Canowindra is, as we have seen, a very special place indeed. Within a year, diggers relocated the spot, earthmoving machinery, trucks and volunteers were dispatched, the rock seam was charted for a hundred meters, and seven tons of Late Devonian fossilized fish, representing thousands of mostly fully articulated individual fossils, were carted off to temporary storage. A volunteer committee of citizens then planned and built a museum, designed exhibits, raised incredible amounts of money, organized events and activities around the theme, and generally responded in classic Canowindra style. The main museum opens this fall (2000) if all goes well.
In most small towns, this Rotary Club talk would have would have been the non-event of the season. But Canowindra is, as we have seen, a very special place indeed. Within a year, diggers relocated the spot, earthmoving machinery, trucks and volunteers were dispatched, the rock seam was charted for a hundred meters, and seven tons of Late Devonian fossilized fish, representing thousands of mostly fully articulated individual fossils, were carted off to temporary storage. A volunteer committee of citizens then planned and built a museum, designed exhibits, raised incredible amounts of money, organized events and activities around the theme, and generally responded in classic Canowindra style. The main museum opens this fall (2000) if all goes well.
In the interim, Alex Ritchie, and particularly his former student, Dr. Zerina Johanson, have been almost as busy. Alone and in collaboration with others, Johanson has rolled out a substantial carpet of papers describing the fish of Canowindra and other Late Devonian sites in the area. Johanson (1997); Johanson (1997a); Johanson (1998); and Johanson & Young (1999). These are not light reading. For the most part, they are part of the painstaking work of species description which is necessary before meaningful questions can be asked about the fossils as living organisms. On the other side, there is much to be said for completely putting aside whatever one does for a day job and immersing completely in minute anatomical detail -- moving slowly between text and the copious illustrations, until the outline of the whole creature gradually emerges from the dense underbrush of detail.
But placoderms are truly obscure. For example, the dermal plates of placoderms are not homologous to the bones of modern fish. In fact, each major tribe of placoderms has its own distinctive pattern of plates, so that they are not even homologous with each other. Thus, for example, an understanding of the paranuchal plates of one antiarch is useful only if one understands how the "neck" plates of other antiarchs fit together. Comparison with other fishes, even other placoderms, is useless. It is, to say the least, a specialized study. Even the jaws and pectoral limbs of placoderms are so unique that they bear little resemblance to other gnathostomes. It is believed that the major jaw adductors of at least some placoderms were medial to the palatoquadrate (i.e. they originated between the upper jaws). The armor-coated pectoral fins of antiarchs in particular are poorly understood and appear at first glance more like the exoskeletal appendages of insects than the fins of other gnathostome fish.
The Canowindra assemblage itself is notably strange. At last count, about
3000 fish were recovered, all in one bedding plane. The vast majority of specimens are from only two species: Bothriolepis yeungae and another antiarch, Remigolepis walkeri, pictured above. In addition, the plane includes a few large sarcopterygians including Canowindra grossi) and an occasional arthrodire Groenlandaspis). The antiarchs are almost all articulated fossils, oriented dorsal-side upwards. However, a few are found disarticulated, mostly in the vicinity of the sarcopterygian remains. Johanson (1997a); Johanson 1998). The size distribution indicates that few were juveniles, enhancing the impression that this death assemblage had perhaps occurred as the result of a single event, perhaps at a time of the year when no young of the year were present.
Johanson (1998) suggests that the assemblage was the result of the drying out of a shallow body of water. Sticking my own paranuchal plates out, I have some modifications, based in part on an entirely speculative view of what those weird Bothriolepis arms were used for. Consider, for example slab at the Canowindra Age of Fishes museum (unfortunately the slab map is no longer posted on the web), as well as the implications of the relatively orderly arrangement. Where the sarcopterygians are found, there are also broken and disarticulated placoderms. As Johanson notes, the big fishes probably thrashed about, as fish out of water will do, and crushed the placoderms in the process. But what about the placoderms? Were they immune from the same reflex? Yet there they lie, rather neatly laid out with scarcely any overlap, perhaps a bit deeper in the slab than the rare sarcopterygians.
Perhaps the antiarchs were not thrashing about because they were already dormant. It has frequently been suggested that the armored pectoral limbs of antiarchs, particularly Bothriolepis, were used to cover the fish with sand as a place of ambush. But the jaws of Bothriolepis yeungae do not even suggest an ambush predator. Johanson (1998). Digging tools have other uses. Dipnoans (lungfish) who inhabit marginal or seasonal fresh water environments will survive dry seasons by digging a hole and becoming dormant -- sometimes for more than a year. If this is true of antiarchs, one might predict a rather uniform posture, minimal overlap, no sign of violent physical movement, fully articulated specimens, and a disproportionate number of burrowing antiarchs -- generally what one sees at Canowindra. In addition, one might predict a near absence of stomach contents and perhaps microstructural details suggesting an absence of ion leaching, insect damage, and rapid drying by comparison with the disarticulated, presumably surface specimens adjacent to the big fish. No one has yet had an opportunity to look at these details. None of this would be conclusive -- almost nothing is in paleontology -- but it might be informative.
The work on the fish of Canowindra has barely begun, and there are probably lifetimes of work and any number of surprises ahead of Johanson and her colleagues. As for the hyperactive inhabitants of Canowindra, who knows? Maybe this time someone at the Rotary Club will decide that what the town really needs is its own starship, or a life-size replica of the Roman Empire...
II. A Brief Overview of the Antiarchs
Even for placoderms, the antiarchs are odd-looking. They are heavily armored; and their eyes, nasal apparatus and pineal are all located in a limited orbital fossa on the dorsal skull. The head itself is literally screwed on backwards. That is, the condyles are on the paranuchal plates of the head and fit into slots on the anterior dorsolateral plates of the body armor. In most placoderms, the reverse is true. That is, the condyles are on the body and the slots are on the head. Antiarch also usually have long, jointed "arms" rather than pectoral fins; and these appendages do not articulate with the body in the normal way. Where it is known, the tail is of a very simple, epicercal type reminiscent of very primitive jawless fish. The ventral surface is flat and simple. It is generally believed that these fishes were bottom-feeders who rarely entered the water column. While one should be properly suspicious of ecological generalities such as this, it is difficult to see any antiarch as an agile, fast-moving predator. Indeed, antiarchs are commonly found in association with arthrodires, suggesting that their niches did not overlap.
One thing we do know about antiarchs is that they probably originated in South China in the Early Silurian, quite likely from arthrodire stock. Young & Zhang (1996). At the time, the South China Province was in sporadic contact with Australia and the rest of East Gondwana to its South. The antiarchs first become numerous in the latest Silurian and Early Devonian of South China and remain more or less restricted to that Province until the Middle Devonian. [1] These early antiarchs were of the yunnanolepidoid type. They were small, but had a large orbital fossa, placed low and far forward on the head. The fossa was surrounded by a depression and/or by ridges. This always included a triangular, medial depression immediately in front of the orbital fossa associated with the small rostral plate (located inside the fossa). The pectoral fin articulation was relatively simple, but rather variable. Janvier (1996). The outline of the fish was low, and thoracic armor relatively shorter than later forms.
Over the course of the Early Devonian, the antiarchs diversified, producing the sinolepids, Minicrania, and other forms. By Middle Devonian times, euantiarchs become common in Australia and Antarctica. At least by antiarch standards, these are evidently fishes with wider horizons. They tend to be larger, with a smaller orbital fossa, placed high and further back on the head. The outline of the fish is taller, suggesting a greater interest in lateral stability in the water column. In some types (asterolepidoids, particularly Remigolepis), the "arms" are secondarily reduced and unjointed, perhaps because the fishes spent more time in open waters and selective pressure for burrowing was less severe. Others (bothriolepidoids) developed the elaborate pectoral articulation characteristic of Bothriolepis. W hile this argues for a bottom-centered existence, it was evidently one which supported a more muscular, higher-energy style of using the pectoral appendages.
Whether or not the euantiarchs remained bottom-feeders, it is clear that they obtained some open-water abilities. By the Late Devonian, the euantiarchs are found world-wide -- a distribution they could hardly accomplish if they were completely unable to navigate in open water. During the Famennian in particular, almost a hundred species of Bothriolepis dominated near-shore environments throughout the world. These, the last of the antiarchs, were frequently quite large, approaching 100 cm if the (usually unknown) unarmored tail is taken into account.
At the end of the Devonian, the antiarchs, as well as all other placoderms, vanished. Not a single placoderm of any type is known from the Carboniferous. Perhaps they became victims of the new tetrapods. Perhaps they were destroyed by climatic changes. Whatever the case, they plainly went from dominance to extinction is a fairly short period of time, by geological standards, along with a number of other near-shore, bottom-feeding groups. ATW
010630
[1] There has been a recent report of antiarch scales in Australia from the Early Devonian. CJ Burrow, CJ & S Turner (1998), Devonian placoderm scales from Australia. J. Vert. Paleontol., 18: 677-695.
References: Carroll 1988); Janvier 1996); Johanson (1997); Johanson 1997a); Johanson (1998); Johanson & Young (1999); Long 1995); Young & Zhang 1996); Zhu & Janvier (1996).
Yunnanolepidoidei = Yunanolepiformes): Yunnanolepis + Chuchinolepis.
$ Head short & much wider than long; triangular preorbital depression present; preorbital recess absent; 'V'-shaped sensory line on postpineal plate; $ lateral plates broad; variable, $ anterior median dorsal plate pointed anteriorly; simple, pectoral fin joints.
References: Janvier 1996); Zhu & Janvier (1996). 010918.
 Yunnanolepidae: Small, very primitive South China antiarchs. Phymolepis, Yunnanolepis.
Yunnanolepidae: Small, very primitive South China antiarchs. Phymolepis, Yunnanolepis.
Yunnanolepidoidei: Chuchinolepidae + *.
Small (<5 cm) forms; head and thoracic armor not fused; premedian plate broad & short; prominent preorbital depression; brachial process absent; no axillary foramen; "wart-like" tubercles; dorsal & ventral thoracic shields almost same length.
Links: Zhu.
References: Janvier 1996); Long (1995); Young & Zhang (1996); Zhu & Janvier (1996).
Note: Phymolepis exhibits two small lateral plates which may be vestigial remnants of the anterior lateral plate of arthrodires. 010924.
Chuchinolepidae: Chuchinolepis (= Procondylepis).
Articular process for pectoral fin unrelated to euantiarch articulation.
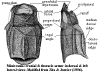 Minicrania: Tiny antiarchs (<2cm) intermediate between yunnanolepidoids & euantiarchs.
Minicrania: Tiny antiarchs (<2cm) intermediate between yunnanolepidoids & euantiarchs.
Squarish skull roof; skull roof and dorsal thoracic shield tightly fused; premedian plate broad & short; very large preorbital depression bounded by prominent ridge; small, crescentic preorbital recess under preorbital depression, just anterior to orbital fossa); orbital fossa relatively far anterior; postpineal plate short & convex; slight dome on postpineal probably myodome for superior oblique eye muscle; short obstantic margin; outlines of 2 semicircular canals visible externally (very large as canals show negative allometry); some additional details of braincase known from internal molds; probably no cartilage between braincase and dorsal dermal bones; sensory line grooves similar to sinolepids and asterolepidoids; but also short 'V' of grooves on postpineal as in yunnanolepidoids; anterior median dorsal plate hexagonal with broad anterior margin; tergal angle (see figure) well forward; median dorsal ridge bracketted by medial dorsolateral ridges; posterior median dorsal plate large and broad; ventral plates also fused, with small, central rhomboidal median ventral plate; ventral plate flat; ventral thoracic plate significantly longer than dorsal plate; "prepectoral corner" tubular, with endoskeletal prepectoral process (as in arthrodires & petalichthyids); simple pectoral joint; plates ornamented with large closely-spaced tubercles.
References: Zhu & Janvier (1996).
Note: among several significant features of this taxon, the co-existence of a preorbital depression and a preorbital recess shows that the two are not homologous. The recess presumably housed the nasal capsules. In yunanolepidoids, the nasal capsules were presumably ventral, not in the depression. 011111.
Sinolepidae: Grenfellaspis, Sinolepis.
lwD-upD of China, Vietnam & Aus.
Antiarcha::: Euantiarcha + *.
Large, squarish heads; preorbital depression present; thoracic armor reduced; large ventral fenestration of thoracic armor; pectoral fin long; pectoral fin articulation may be primitive version of euantiarch articulation, including funnel pit with "remnant" of scapulocoracoid and axillary foramen.
References: Janvier 1996); Long (1995); Zhu & Janvier (1996).
Note: possibly paraphyletic (i.e. ancestral to euantiarchs). 010624.
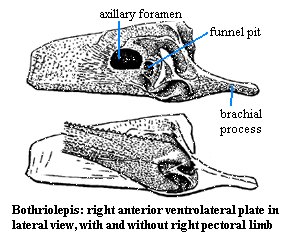 Euantiarcha:
Euantiarcha:
Rnge: lwD-upD.
Relatively uniform morphology; $ short postorbital skull; $ helmet-shaped, ball-in-socket pectoral joint; funnel pit with "remnant" of scapulocoracoid; axillary foramen for pectoral nerves and circulation.
Image: modified from Janvier (1986: 165 fig. 4.52 F-G). 010626
Microbrachiidae: Microbrachius. Tiny (<2 cm) antiarchs transitional between Yunnanolepidoids and Bothriolepidoids.
Preorbital depression present.
References: Janvier 1996). ATW020806.
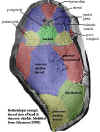 Bothriolepidoidei: Bothriolepis.
Bothriolepidoidei: Bothriolepis.
mD-upD (or lwC) worldwide.
Preorbital depression absent; separate rhinocapsular recess (i.e. a separate nasal capsule compartment under the premedian plate); unpaired semilunar plate.
Links: Bothriolepis sp.; UALVP
1835 Bothriolepis canadensis replica; L'ère des Poissons; Class Placodermi
Upper Image: This is the very common "squashed bug" view of Bothriolepis. This is how it is found in the field; but, bear in mind that in life, the lateral plates would have been much more lateral, i.e. vertically oriented. You wouldn't look so trim either, after 300 My under several tons of rock.
Lower Image: Bothriolepis is arguably the most common middle and Late Devonian fish. Bothriolepids became very large and acheived worldwide distribution in the up (Frasnian & Famennian). One of the most widely-recognized of all the ancient Armoured fishes, this is Bothriolepis. The box-like body was enclosed in armor plates, and provided armoured protection from predators. Bothriolepis was the most successful of all the placoderms, with some 100 species known from every continent, including Antarctica.
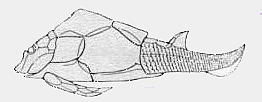 Asterolepidoidei: Asterolepis, Pterichthyodes, Remigolepis.
Asterolepidoidei: Asterolepis, Pterichthyodes, Remigolepis.
Preorbital depression absent; posterolaterally expanded postmarginal plate; narrow lateral plate; short nuchal plate; very long thoracic armor; pectoral appendage loses distal joint in Remigolepis.
Links: Volume 16 (V2).
Image: Remigolepis. ATW010626.