 Holocephali
-- Basic Anatomy
Holocephali
-- Basic Anatomy| Chondrichthyes | ||
| The Vertebrates | Holocephali |
| Vertebrates Home | Vertebrate | Vertebrate |
|
Abbreviated Dendrogram
Thelodonti
├─Placodermi
│
└─Eugnathostomata
├─Chondrichthyes
│ ├─Eugeneodontida
│ └─Chondrichthyes (Crown)
│ ├─┬─Cladoselachida
│ │ └─Symmoriida
│ │ ├─Symmoriidae
│ │ └─Holocephali
│ │ ├─Cochliodontidae
│ │ └─┬─Menaspidae
│ │ └─Chimaeriformes
│ └─Elasmobranchii
│ ├─Xenacanthida
│ └─Ctenacanthiformes
│ ├─Ctenacanthidae
│ └─Euselachii
│ ├─Hybodontiformes
│ └─┬─Synechodontiformes
│ └─Neoselachii
└─Teleostomi
|
Contents
Overview |
The group here considered used to be called the "Bradydonts" and was said to be characterized by a low rate of tooth replacement. The animals usually comprehended by this term turned out to be a broad selection of unrelated basal chondrichthyans. Thus, the term "Holocephali" or "Holocephalomorphi" (the distinction, if any, has escaped us in the literature) is preferred for the group including the living chimaeroids and their Mesozoic cousins. However, as a descriptive term, "Holocephali" is useless. Almost all the bradydont genera are also holocephalic. That is, the palatoquadrate is fused to the braincase.
This whole area of phylospace is badly in need of revision, since no one has done a serious cladistic analysis. However, there are no doubt industrious graduate students out there somewhere sitting on the requisite data. In due course, we will see what it shows. The story is an important one, since the chimaeras are the most primitive of all living gnathostomes. As discussed below, an understanding of their evolution would do a good deal to working out the intractable jaw story.
Holocephali
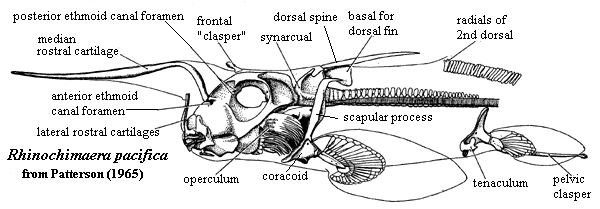
-- Basic AnatomyThe figure on the left shows one of the living chimaeroids, Rhinochimaera pacifica. It is hard to say what a "typical" holocephalan might be, since the Holocephali are rather variable and poorly known, as well as being downright weird. Rhinochimaera is probably not typical of anything. However, it will do as a means to introduce some of the unique features of the Holocephali, as well as the living members of the Chimaeroidei.
One of the most obvious features is that the creature looks more like a grasshopper than a fish. There are several reasons for this misleading gestalt impression. The most obvious is the flamboyant display of "labial cartilages," including the median rostral cartilage that looks like an antenna in Rhinochimaera. During development these cartilages look suspiciously like gill arches, and -- in a few cases -- have similar segmentation. Whether the labial cartilages are actually related to gill arches and, if so, whether they are neomorphic duplications or throw-backs to a premandibular arch, is anyone's guess. In addition to these cartilages, holocephalans may bear a frontal "clasper," a structure which appears to be derived from a fin spine and which bears ornamentation suggestive of the symmoriid head dress. Very likely, that's exactly what it is.
The jaw suspension is, of course, holocephalic. That is, the palatoquadrate is fused to the braincase. On this subject, more anon. However, note that the postorbital braincase is extremely slight. The gill arches are largely tucked in below the posterior braincase and are covered with an operculum. The holocephalan operculum is probably not homologous with the operculum of Teleostomi, and is supported by the hyoid arch. Not only is the holocephalan structure different, and somewhat differently placed, but some Acanthodii lack an operculum, suggesting that the teleostome operculum developed after that line diverged from the sharks.
Like their symmoriid cousins and/or ancestors, the holocephalans developed a rather complex relationship between the pectoral appendages, as well as between the pectoral girdle and the first dorsal fin. Many holocephalans have coracoids fused across the ventral midline. In addition, the scapulae may be closely associated with each other and with the basal cartilage(s) of the first dorsal. The anterior arcualia of the notochord are also fused into a synarcual structure which develops a strong dorsal process, probably as a consequence of fusion with an exapted dorsal fin basal. The first dorsal fin spine (there is never a second dorsal spine) articulates with the synarcual, which enables the spine to swivel up and or lie flat. Lund 1977).
The notochord posterior to the synarcual is nominally "free." However, like many freedoms, one has to look at the fine print. The notochord is actually constrained by numerous thin rings of perichondral calcifications. These are arranged without any consistent relationship to the expected chordal or muscular "segments," an arrangement referred to as polyspondyly.
Finally, note that the pelvic girdle (which is not fused) bears -- not only the usual posterior myxipterygial clasper -- but also an anterior structure referred to as a tenaculum. The apparently also has some role in copulation, display or courtship. The living chimaeroids exhibit marked sexual dimorphism. This dimorphism is not limited to sex-specific structures, such as the male tenaculum, but also in the shape and size of other structures. Males are generally smaller, more gracile and more angular than females.
 Turning to Teeth
Turning to TeethFinally, we must turn, albeit with a deep sigh, to the subject of dentition. Let us be frank. For the most part, the literature relating to tooth replacement is hopelessly boring, as well as confused. The skeptical reader may question our experience, even our dedication, to the cause of condrichthyo-dentistry. But even the late Dr. Colin Patterson [1], after a lifetime of looking gift sharks in the mouth, was forced to admit that, "the nomenclature used for the various surfaces of the tooth plates in chimaeroids has been varied and so inconsistent as to be chaotic." Patterson 1992: 37).
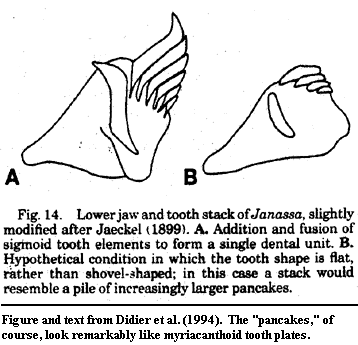
Out of this bewildering tangle, we are fortunate to have extracted a few points of clarity. In particular, Didier et al. (1994) is a paper with an all-star cast of authors and a very firm conclusion: the tooth plates of chimaeriforms are ordinary chondrichthyan dentition. They are neither unique structures nor placoderm-like gnathal plates. Each chimaeriform tooth plate represents a single tooth family, usually with two members which grow throughout the life of the animal in a perfectly normal shark-like way. In the living chimaeras, Callorhinchus and Chimaera, the authors were able to show the persisting boundaries between the individual teeth. In extinct chimaeriforms, they identify similar structures, and derive the nature of the probable intermediates and variants yet to be discovered. Such complete answers -- particularly with clearly testable conclusions -- are relatively unusual in paleontology, and doubly rare in world of Paleozoic sharks.
Thus, far from being placoderm-like, the tooth plates of chimaeras are at least distantly reminiscent of the ubiquitous palatal "fang pairs" of the Sarcopterygii and their strange offspring, the tetrapods! Sadly, it is unlikely that this research will be extended. The Didier group used a process they term "artificial petrification." This method seems to produce distortion-free thin sections of startling detail and clarity. However it involves a drying step which takes eight to ten months -- scarcely faster than natural petrification compared to the usual academic publication cycle.
 Holostyly: the Suspension Mounts ...
Holostyly: the Suspension Mounts ...Having disposed of general anatomy and special dentistry, we are now positioned to look at the real stuff: the jaw suspension. After all, few people (Bashford Dean, perhaps, excepted) study ratfish for their intrinsic beauty, for spiritual inspiration, or for the millions undoubtedly to be made peering at dried tritors. No, it is only the jaw that matters. Chimaeroids are the most primitive of living gnathostomes. To find the plesiomorphic state of the holocephalan jaw is, just possibly, to find the primordial state of all jaws. And, from there, it is but one step to the Holy Grail of vertebrate paleontology -- the origin of jaws.
A few years ago, the intrepid knightly band of Eileen Grogan, Richard Lund and Dominique Didier (Grogan et al., 1999) may, like Lancelot, have partially completed the Great Quest for the this Grail, or at least have seen a vision of it, "All palled in crimson samite, and around / Great angels, awful shapes, and wings and eyes." Well -- perhaps not the bit about the angels and wings. But the eyes and, definitely, most definitely, the awful shapes. The included smeary photomicrographs are certainly that. But, like Lancelot or Grogan), we may see aspects of the Great Truth:
... through a stormy glare, a heat
As from a seventimes-heated furnace, I,
Blasted and burnt, and blinded as I was,
With such a fierceness that I swooned away--
O, yet methought I saw the Holy Grail ...
The issue goes back to the Hundred-Years' War over the ancestry of holocephalan relationships. Clearly, those favoring a placoderm ancestry expected an unbroken lineage of holostyly. Those contending for a selachian (shark) origin expected a gradual conversion to chimaeran holostyly from selachian hyostyly, the typical shark condition in which the jaw suspension is provided, at least in part, by the hyomandibular. For some time, the evil Skandic forces of Stensiö, Ørvig, Bendix-Almgreen, and other placoderm partisans seemed to carry all before them. It was repeatedly shown that the hyoid arch showed no traces of involvement -- past or present -- in the chimaeroid jaw suspension. The chimaeran hyoid is, it turns out, quite happy supporting the operculum and has no interest in the palatoquadrate. Patterson 1965). Worse, there is no trace of anything resembling an otic articulation for the palatoquadrate. The otic articulation is common in both modern and Paleozoic sharks and was seen as a probable intermediate between a free palatoquadrate (hyostyly) and one fused to the braincase (holostyly). As it turns out, chimaeroids scarcely have a recognizable otic region, much less an otic jaw articulation.
For a number of reasons extraneous to the present discussion, the placoderm connection began to look less viable by the 1960's. However, this central difficulty with the jaw suspension remained. Grogan et al. ultimately solved this problem by re-examining the embryology of the chimaeroid jaw, a subject not addressed since Dean 1906), in the new light of new specimens of putatively intermediate forms from the Early Carboniferous Bear Gulch chondrichthyan fauna. They found that the primitive state for all chondrichthyans (and quite likely placoderms as well) is autodiastyly. In this condition, the palatoquadrate is suspended from the braincase in two places (a) an anterior articulation from the ethmoid and (b) a posterior articulation with the basitrabecular. The latter structure requires explanation. The reader may recall that the vertebrate skull is derived from two pairs of cartilagenous rods: the anterior trabeculars and the posterior parachordals. See, diagram from Braincase : Overview. The basitrabecular is a derivative of the polar cartilages at the intersection of the trabeculars and the parachordals. From this version of autodiastyly it is straightforward to derive both holo- and hyostyly, as well as other variations on those themes. Either the palatoquadrate fuses with the base of the braincase for support as it grows (holostyly), or it enlists the aid of other structures such as the otic capsule or the hyomandibular (autostyly, hyostyly).
This understanding is particularly interesting in that it places the plesiomorphic jaw suspension for sharks at the same place as the basicranial jaw articulation for sarcopterygians. This strongly suggests that Grogan et al. have arrived at a really fundamental truth. The question which now needs an answer is whether the basipterygoid process of sarcopterygians and tetrapods is homologous with the basitrabecular process of sharks. That is, does the basipterygoid also form with the participation of the polar cartilages at the posterior end of the trabeculae? The figures in Romer (1956) suggest that this is so. Quite likely, the answer is already known from the musty old comparative embryology treatises of deBeer and Gaupp, all of which were written about the same time as Dean's work in 1906.
Notes: [1] Patterson (1933-1998) is one of our personal intellectual heroes. He was not always right, but his views were always carefully considered and researched, balanced, objective and expressed in a prose which was clear and direct. For a fact, it is remarkable how many of the dead shark workers are, and have always been, scientists of powerful intellect and remarkable originality. This is all the more remarkable because the material is horribly scrappy and rare; the work, difficult and utterly without public acclaim.
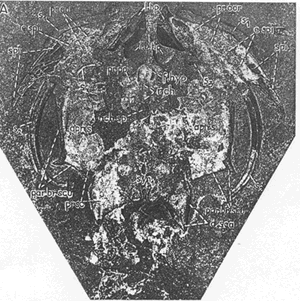
 Menaspidae
MenaspidaeThe reconstruction shown here -- and in many other places -- is the reconstruction of Svend Bendix-Almgreen 1971) who asserted that Menaspis was a placoderm. Slightly earlier, Colin Patterson had published a rather different reconstruction. In Patterson's view, the orbits were located much further back, and the various protuberances were located on a much longer and thinner rostrum. The choice of Bendix-Almgreen's illustration below is due largely to the vagaries of availability. Many libraries carry Lethaia. Few have the Proceedings of the Fourth Nobel Symposium. Truth be told, neither model is particularly convincing, simply because the fossil material is so opaque and ambiguous.
There are also other possibilities. For example, Bendix-Almgreen illustrates some of his points with pictures of Recent rays. These are almost too convincing. Could it be that the mysterious head spines (apparently labial cartilages gone berserk) actually supported a ray-like extension of the pectoral fin all the way to the anterior margin of the animal? That is our personal favorite, but that model is at least as speculative as all the others.
Holocephali: (= Bradydonti?) Harpagofututor, Ischyodus
fr mD.
Phylogeny: Falcatidae: Damocles + *: Cochliodontidae + (Menaspidae + Chimaeriformes).
$ precerebral fontanelle absent reversal) [CS01*]; $ palatoquadrate fused with braincase holostylic) [CS01*]; $ loss of expanded palatoquadrate otic process (reversal) ($ and corresponding otic articular fossa) [CS01*]; soft tissue operculum [L77]; primitively 3 tooth positions in each jaw? [L77]; 2 tooth plates upper jaw, 1 in lower; teeth not replaced; spiracle absent in adult; operculum present; postorbital articular process for palatoquadrate absent (reversal) [CS01*]; anterior vertebrae fused with dorsal spine; $ loss of anteriorly concave (lateral view) spine shape reversal) [CS01*]; loss of triangular cartilage from posterior dorsal fin reversal) [CS01*]; reduced caudal fin; most anterior pectoral radial largest [CS01*]; mixopterygial claspers, as sharks [DD93]; tessellate perichondral prismatic calcification of cartilage, as sharks [DD93] [L77]; sensory lines in open grooves, with crescentic "ring scales" [L77] (stating that it is plesiomorphic for sharks); scales with neck canals, as sharks [DD93] strong sexual dimorphism in virtually all species [L90].
Coates & Sequeira (2001) [CS01]; Duffin & Delsate (1993) [DD93]; Lund 1977) [L77]; Lund (1990) [L90].
Notes: [1] "[CS01*]" indicates synapomorphies which were calculated from the data given in Coates & Sequeira (2001) using very different software (PHYLIP). Using the full Coates & Sequeira data set, this analysis yields a single most parsimonious tree identical to that found by the authors, but requires 3 more steps to reach it. The actual data used to generate these synapomorphies is slightly different and gives slightly different results. [2] All these reversals are not as unlikely as it may appear. They merely reflect the increasing consolidation of the palatoquadrate with the braincase and concomitant adaptation to a durophagous, slow-moving, benthic habit -- a tendency finding its logical endpoint in the holocephalans. ATW011209.
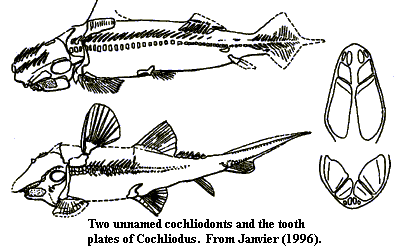 Cochliodontidae: Sometimes spelled "Cochleodontidae." Cochliodus, Cranodus, Crassidonta, Cymatodus, Deltodus, Dichelodus, Diplacodus, Erismacanthus, Helodopsis, Icanodus, Macrodontacanthus, Menaspacanthus, Poecilodus, Psephodus, Synthetodus, Thoralodus, Xenodus.
Cochliodontidae: Sometimes spelled "Cochleodontidae." Cochliodus, Cranodus, Crassidonta, Cymatodus, Deltodus, Dichelodus, Diplacodus, Erismacanthus, Helodopsis, Icanodus, Macrodontacanthus, Menaspacanthus, Poecilodus, Psephodus, Synthetodus, Thoralodus, Xenodus.
Late Devonian [C88] to Late Permian of North America, Europe, Australia, & South Asia
Phylogeny: Holocephali: (Menaspidae + Chimaeriformes) + *.
Characters: largely a tooth plate taxon. Bodies somewhat ventrally flattened, but with notable variety of body forms [L90]; paired mandibular and occipital spines [L90]; paired frontal spines with "brush" complexes (but paired and presumably derived from labial cartilages, not fin spine) [J96]; "bradydonts" having teeth with curved or rounded surfaces, usually with folds or grooves; multiple small anterior tooth plates in upper jaw [C88]; dentition with complex hypermineralized tissue arching out into jaw [D+94]; dentition typically (Cochliodus) anterior tooth family plus two posterior plate pairs on both jaws [L77]; at least one species (Synthetodus) has median symphysial plates, possibly in both jaws [L77]; anterior teeth may be individual, whorls, or small plates [L90]; branchial arches do not always lie under the neurocranium in most Holocephali [L77]; most with conventional holocephalan anterior dorsal fin, but some may lack synarcual and all anterior dorsal fin structures [L90]; second dorsal fin supported on basal [L90]; .
Links: Cochliodontidae; coch.jpg (weird teeth); kocochli1.htm; Deltodus sp - Mississippian Crusher Shark Tooth - Salem Formation; Deltodus; Moscow.htm; Marine Fossils from the Pennsylvanian Deposits of Ferguson, MO.; shark; matrix; Sharks and Shark-like Creatures of the St. Louis Limestone, St. Louis, Mo.; Indiana.htm; Aspects of enameloid ontogeny in fossil and recent selachians with comments on enameloids and their occurrence during early lower vertebrate; Dinosaurclass.Com; Dossier sur les elamobranches.
References: Carroll (1988) [C88]; Didier et al. (1994) [D+94]; Janvier (1996) [J96]; Lund 1977) [L77]; Lund (1990) [L90]. ATW020905
 Menaspidae: Deltoptychius, Menaspis.
Menaspidae: Deltoptychius, Menaspis.
Range: Early Carboniferous to Late Permian of North America & Europe.
Phylogeny: Holocephali:: Chimaeriformes + *.
Characters: mouth terminal [B71]; head not laterally compressed [P65]; rostrum not enlarged [P65]; nares probably dorsal or anterodorsal [B71]; two large pairs of sickle-shaped (frontal?) cephalic spines anterior to orbits [B71]; frontal spines of "dense trabecular dentine" [Z81]; several large dermal plates on skull roof and lateral skull [P65]; orbits dorsal and close together [B71]; 1 small pair of sickle-shaped cephalic spines posterior to orbits [B71]; (posterolaterally- directed?) postorbital process [B71]; frontal claspers very reduced or absent [P65]; somewhat rectangular otic capsules [B71]; jaw suspension holostylic [B71]; jaw at a distinct angle to body axis, with anterior jaw far dorsal to jaw articulation in closed position [B71]; rami of lower jaw diverge at about 90° [B71]; tooth plates thin & without tritors [P65]; 1 lower pair of tooth plates [P65]; symphysial teeth on lower jaw [Z81]; dentition close to chimaeroid model, but uncertain; pair of robust, relatively straight ornamented bony spines extending almost laterally from anterior of coracoid as prepectoral spine Bendix-Almgreen) or from lower jaw as mandibular spine (Patterson) [B71] [P65]; "prepectoral" spine continuous with coracoid bar [B71]; gill openings probably dorsolateral [B71]; possibly two synarcuals or 2-part synarcual [B71]; notochord without calcified rings [P65]; anterior dorsal fin spine absent [P65]; anterior dorsal fin apparently absent [P65]; coracoid located far anterior of usual position [B71]; scapulocoracoid coracoid processes fuse anteriorly as coracoid bar and floor the branchial chamber ventrolaterally and posterolaterally [B71]; scapular processes may fuse or articulate with a synarcual [B71]; dermoscapular spine associated with a posteriorly directed process of scapular region [2] [B71]; possible complete exoskeletal analog to the shoulder girdle [B71]; pectoral fin with three basals & 19 radials [B71]; complex pattern of dermal scutes [B71]; scutes coalescing to form dermal armor [C88]; two rows of enlarged scales on dorsal trunk [P65]; scales cyclomorial (!?) [P65].
 Notes:
[1] Bendix-Almgreen's article is devoted to discussion of the difference between his model and the model of Patterson discussed above. Accordingly the notes from [B71] are precisely those points which were -- and remain -- controversial. [2] this spine looks displaced to us [3] Zangerl (1981) reports that additional material recovered after the Bendix-Almgreen (1971) paper confirms Patterson's understanding of the pectoral girdle, and everything else except the position of the orbits, in which Bendix-Almgreen seems to have been generally correct..
Notes:
[1] Bendix-Almgreen's article is devoted to discussion of the difference between his model and the model of Patterson discussed above. Accordingly the notes from [B71] are precisely those points which were -- and remain -- controversial. [2] this spine looks displaced to us [3] Zangerl (1981) reports that additional material recovered after the Bendix-Almgreen (1971) paper confirms Patterson's understanding of the pectoral girdle, and everything else except the position of the orbits, in which Bendix-Almgreen seems to have been generally correct..
Image: (right) the "Berlin specimen" from Bendix-Almgreen 1971). See also life construction in text, supra.
Links: Ortlam, D.: Neue Aspekte zur Deutung von Menaspis armata Ewald (Kupferschiefer, Zechstein 1, Deutschland) mit Hilfe der stereoskopischen Röntgentechnik. ISBN ES186008100; Dossier sur les elamobranches.
References: Bendix-Almgreen 1971) [B71]; Carroll (1988) [C88]; Patterson (1965) [P65]; Zangerl 1981) [Z81]. ATW030501.
checked ATW030924
{kind=link}
{kind=link}