 Esociformes: Not As Plain as a Pikestaff
Esociformes: Not As Plain as a Pikestaff| Euteleostei | ||
| The Vertebrates | Neognathi |
| Vertebrates Home | Vertebrate | Vertebrate |
|
Abbreviated Dendrogram
Teleostei
│
└─Euteleostei
├─Ostariophysi
│ ├─Anotophysi
│ └─Otophysi
│ ├─Cypriniformes
│ └─Siluriphysi
└─Neognathi
├─┬─Salmoniformes
│ └─Esociformes
│ ├─Umbra
│ └─┬─Dallia
│ └─┬─Novumbra
│ └─Esocidae
└─┬─Argentiniformes
└─Neoteleostei
├─Stenopterygii
└─Eurypterygii
├─Cyclosquamata
└─Ctenosquamata
├─Scopelomorpha
└─Acanthomorpha
|
Contents
Overview |
Esociformes: Not As Plain as a PikestaffFor a group with only 4 living genera and a minimal number of complicating fossils, the relationships of the Esociformes -- both internal and external -- are absurdly difficult. Briefly, this is the domain of two freshwater clans, the pike and the mudminnows. The traditional classification, which may be correct, places the pike (genus Esox) into one family (Esocidae) and the mudminnows into another ("Umbridae"). Esox is a complicated genus of big (up to 150 cm), visually- oriented predators with big appetites, broad tastes, and nasty dispositions. The various species are found in cold to cool freshwater habitats all over the northerly parts of the Northern hemisphere, where they are much prized as sport fish.
The classical Umbridae include three genera of odd and disjunct distribution: Novumbra, Dallia and Umbra. Novumbra is the so-called Olympic Mudminnow; although, in fact, it is not notably athletic, nor even a sport fish. Rather, it is a very small (<8 cm) and undistinguished fish restricted to a few small watersheds around the Olympic Peninsula of Washington State in the northwestern United States. Dallia is neither minnow-like nor fond of mud. Its common name is the Alaskan Blackfish, and it looks rather more like a smallish (30 cm) pike with big, fan-like fins. There are either one or three species, depending on whom you believe, distributed in Alaska, the islands of the Bering Sea, and northeastern Siberia. Finally, Umbra includes three species of small insect eaters, two of which are quite common in the central and eastern parts of Canada and the northern U.S., with a third species now restricted to odd sections of the Danube Basin in Europe.
Just to make things deceptively simpler, we note that almost everyone agrees on the relative positions of the genera. Esox and Umbra are the bookends, and Novumbra is either closer to Esox or further from Umbra, than is Dallia. This position is not completely without dissenters. Kettler & Whitt (1986) propose a Novumbra-Dallia clade, with both closer to Umbra than to Esox, based on the tissue distribution of lactate dehydrogenase isozymes. However, as we will see, there are excellent reasons for deep suspicion of any phylogenies based on metabolic enzymes in this corner of phylospace. Briefly, it appears that almost all of the esociforms were subject to severe selection for cold and drought tolerance in the recent geologic past, which can do very odd things to structures of metabolic proteins. For the same reasons, we may ignore the cytochrome b mtDNA sequence study of López et al. 2000). López quite correctly rejects these results himself because of the overwhelming evidence of mutational saturation. That is, the same sites have mutated so many times that the codon at many positions is essentially random and carries no phylogenetic signal. The tests he applies are elegant and worth of emulation by others in the molecular crowd.
| Euteleostei
├─Esox
└─┬─Novumbra
└─┬─Dallia
└─Umbra |
Euteleostei
├─┬─Novumbra
│ └─Esox
└─┬─Dallia
└─Umbra |
Euteleostei
├─Umbra
└─┬─Dallia
└─┬─Novumbra
└─Esox |
|
In any case, the problem is not branch order, but the location of the root. The agreed branch order is compatible with three different arrangements, as shown in the table, depending on where the root falls. The traditional arrangement -- which we have recently abandoned -- is at left of the table. It has been tested cladistically only once that we know of, by Wilson & Veilleux (1982). This study was, obviously, performed over 20 years ago, right at the beginning of the cladistics era. It is not at all clear that the results would be the same if performed today. Two observations will suffice to make the point.
First, Wilson & Veilleux (1982) is one of the best works of comparative osteology we have ever seen for any taxon, with one glaring exception. The osteology of Esox itself was not reviewed, and none has been published since the 1800s. Nor did Wilson & Veilleux examine the two excellent fossils in the area, Palaeoesox and E. tiemanni. The former was not available to Western workers at the time, and the latter had not yet been fully described (Wilson completed this work a few years later).
Further, the polarity of the characters used by Wilson & Veilleux is suspect because the position of esociforms within the teleosts was unknown. At the time, it was believed that Esociforms (usually referred to as "Esocoidei" in the older literature), were either a kind of salmoniform as assumed by Wilson & Veilleux), or were even more basal -- perhaps even the sister of Euteleostei. Opinions on this issue seemed to be changing by about 1990, when two, large-scale cladistic studies were published by Begle in 1991 and 1992. These appeared to affirm the traditional view. Begle abruptly left ichthyology in 1993 for reasons which were unclear. It came as something of a shock when, several years later, Johnson & Patterson 1997) published strong evidence that Begle's work was corrupted by gross carelessness and, very probably, scientific fraud. Their reanalysis of the materials placed Esociformes as the sister of Neoteleostei. This conclusion, as Johnson & Patterson stated, was very weakly supported, but made reasonably good intuitive sense. If even partially correct, the character polarities assumed by Wilson & Veilleux in 1982 were well off the mark.
This brings us back to the work of López et al. (2000). In addition to their study using cytochrome b mtDNA, these workers also studied the evolution of the 16S rRNA gene in esociform mitochondria. Let us be frank. We are not friends of sequence-based molecular phylogenies. The reasons for our deep suspicion are somewhat beyond the scope of this essay, but may be summed up by noting that the primary structure of biomolecules is only related at third hand to their function; while natural selection -- by definition -- occurs at a functional level. Still, not all evolution is about natural selection, and much has been made about the role of chaotic and random events at the DNA level.
In any case, López et al. (2000) was a reasonably good study, all the more impressive because the tests used were able to detect that the cytochrome b portion of the project was not useful. Strangely enough, so was the rRNA portion when osmerids (smelt) were used as the outgroup. However, when salmonids were used, the results were very clean. This is odd, since the salmonids and osmerids were supposed to be sisters; and even odder because it resulted in both Dallia and Novumbra being closer to Esox than to Umbra. López et al. noted that, when other taxa were used as outgroups, the result was an unresolved polytomy among the esociform genera. Finally, the 2000 study raised some methodological questions. All things considered, it was a fairly simple matter to apply our usual techniques of arm-waving and obnoxious rhetoric and dismiss the whole thing -- which is exactly what we did.
Sadly, for those of us who have labored many years to perfect the high arts of arm-waving and obnoxious rhetoric, science has a disgraceful way of overcoming even the most refined bombast with fact. And so we are -- once again -- pained to report the destruction of the subtle and delicate products of human creativity under the crushing weight of soulless data.
López et al. (2004), a sequel to the 2000 study, is a convincing paper on a number of levels. This is novel, since all other studies of Esociform taxonomy have either had serious limitations, or have yielded only weakly supported results. It is a molecular study, but discusses and deals with morphological information in a reasonable way. It uses a really large and representative sample of lower teleost species. Part of the work uses mitochondrial DNA. However, the usual questions about mtDNA need not detain us because the López group gets the same result with a long nuclear sequence from RAG-1 (recombination activation gene 1).
| Neognathi
├─┬─Salmoniformes
│ └─Esociformes
│ ├─Umbra
│ └─┬─Dallia
│ └─┬─Novumbra
│ └─Esocidae
├─"Osmeroids"
└─┬─Argentiniformes
└─Neoteleostei |
| "Osmeroids" is a placeholder for two osmeriform groups, the Osmeroidea (smelt) and Galaxoidea. The Galaxoidea, in turn, are paraphyletic and contain the Stomiiformes lanternfishes) which are often regarded as part of the Stenopterygii. |
Unlike many genes used in phylogenetic studies, RAG-1 is a really good proxy for long-term vertebrate evolution. RAG-1 has a number of indispensable properties. In fact, it is almost unique in having all of these properties at once. (1) It is a single copy gene with no messy allelic variations. Even more importantly, (2) it was acquired by the vertebrates through lateral gene transfer from a viral genome about the same time as the first vertebrates evolved. This is critical because it means that there are no close molecular relatives in the vertebrate genome from which RAG-1 might have evolved, and with which it might be confused. There are no analogs, paralogs, orthologs, or other firewood to confuse us. (3) RAG-1 is used by vertebrates only as part of the machinery which creates somatic gene variability for the immune system, a system which is rather indifferent to the organism's ecological role or environmental specializations. (4) RAG-1's only substrate is DNA, and (5) the gene product of RAG-1 has no other function. Thus, since DNA is always chemically the same, and RAG-1 interacts only with one substrate for one reaction with one purpose, it is not subject to much selective pressure by ecologically-controlled factors, variable substrates, diversity of metabolic pathways, or other forces which might produce either homoplasy or adaptive divergence. (6) It is extremely slow to evolve -- particularly the portion of the gene used by the Lopez group -- so that saturation concerns are minimal. Finally (7), the gene normally contains no introns, thus reducing concerns that RAG-1 genes in closely related organisms may diverge quickly based on selection for interaction with some rapidly-evolving RNA editing enzyme. Our criterion #7 is subject to some qualification since it turns out that 1-2 small introns have evolved, uniquely, in certain teleost fishes. However, this is unlikely to have caused major errors, given the results of the 2004 study.
In short, RAG-1 is about as bullet-proof as any DNA sequence can be. Certainly it has resisted all of our efforts to shoot holes in it. In fact, we were left feeling like one of those hapless villains in an old Superman comic. After the Master Criminal has exhausted all of his reserves of evil design, his legion of homicidal minions, and a vast arsenal of high explosives, there comes a moment of pregnant silence. Then, as the smoke clears, we behold the Man of Steel standing yet unharmed, hands on his hips and the customary superior smirk on his face -- that kind of feeling. At this point, the usual formula requires the Son of Jor-El to utter some insipid and utterly gratuitous moralism as the frustrated Megalomaniac of the Month seethes with dumbfounded rage. Fortunately, we are able to avoid such a degrading denouement to the destruction of our own fiendish phylogenetic plots, by simply presenting the López et al. (2004) revised cladogram (in highly simplified form) in the table.
It appears that neither the Umbridae (Umbra + Dallia + Novumbra) nor the Protacanthopterygii Salmoniforms + Argentiniforms) [1] are valid taxa. Umbra, not Esox, is the more plesiomorphic genus. The root of the Esociformes lies between Umbra and Dallia, rather than between Esox and Novumbra. Pikes are closely related to the salmonids after all, but the argentiniforms are somewhat more distantly related. One of the more attractive parts of the 2004 scheme lies in in its heterodox grouping of "osmeroids," a group explained in the table. The lanternfishes of the Stomiiformes look a great deal like mutant smelt, and that is just where they fall out in the cladogram. At the moment, our own coverage of this region of phylospace is too superficial to take this arrangement properly into account. We look forward to further development in this area.
ATW050604. Public domain. No rights reserved.
Update: About eight weeks after we wrote the last revision in 2005, a group of fish systematists met in Madrid, at which meeting Newbrey et al. 2005) came to similar conclusions regarding the arrangement of genera within the esociformes. It seems that scales and vertebrae of Cretaceous esociforms are now common enough to do some statistical work. The data support the idea that the basal esociform groundplan was similar to that of the umbrids, to the extent that they were all small fishes, with relatively small short lifespans and slow growth rates. The modern pikes diverged from this condition during the Paleocene.
ATW080122. Public domain. No rights reserved.
[1] As originally conceived, Protacanthopterygii included both Salmoniformes and Esociformes. Although the position of this group has changed, perhaps that would be an appropriate name for the salmonids and pikes, as currently arranged.
Range: from the middle Cretaceous
Phylogeny: Euteleostei :: + * : (Salmoniformes + Esociformes) + Argentiniformes + Neoteleostei).
Characters: depressible teeth present [$JP97]; uroneural 3 absent [$JP97]; cheek & operculum with scales [$JP97]; skeletal bone acellular [$?JP97].
References: Johnson & Patterson (1997) [JP97]. ATW 080122.
 Salmoniformes: salmon (Salmo). Salmonoids.
Salmoniformes: salmon (Salmo). Salmonoids.
from the Early Cretaceous
Neognathi :: Esociformes + *.
Characters: Maximum length up to 1.5 m; single supramaxilla [$?JP97]; dermethmoid & supraethmoid not fused [$JP97]; posttemporal fossa not roofed [$JP97]; deep posterior myodome with eye musculature passing through and attaching to trunk muscles; on pharyngobranchials 4-5, only 5 has toothplate [$JP97]; 7-20 branchiostegal rays; vertebrae 50-75; epineurals fused on 0-1 vertebrae [$JP97]; epipleurals not ossified [$JP97]; adipose fin present; last few neural & hemal spines, distally ends expanded anteroposteriorly [$JP97]; uroneural 2 broad & anterodorsal to uroneural 1 [$JP97]; mesocoracoid present; gill membranes extending far forward, free from isthmus; pelvic axillary process present; last three vertebrae turned up; scales lack radii [$JP97]; nuptial tubercles present [$?JP97]; tetraploid karyotype; parr marks in young of most species; diadromous [$JP97].
Image: Salmonid from FishBase WWW:Taxonomy (former page).
Links: IWR: Taxa: Salmoniformes; Salmoniformes.
References: Johnson & Patterson (1997) [JP97]. ATW080122.
Esociformes: pikes & mudminnows. Coextensive with Esocoidei. The latter name was used when this taxon was believed to be part of Salmoniformes. †Boltyschia, †Palaeoesox?
Range: from Late Cretaceous (Campanian), currently restricted to North America, Russia & Europe.
Characters-1: The following characters were assembled largely from sources which conceived of Esox as being primitive for the Esociformes, and therefore is somewhat suspect as a list of esociform characters. However, except for polarity problems, it still has utility.
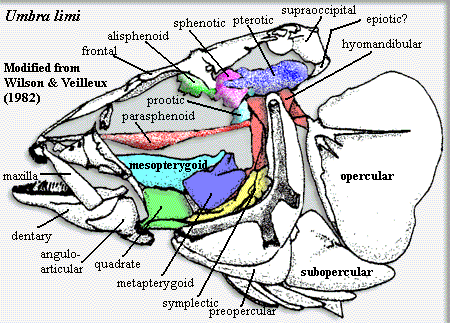
![Esox mandible from [J75]](images/EsoxMandible.gif) Characters: maxilla toothless but included in gape [N94]; single supramaxilla [$?JP97]; median unpaired supraoccipital with posterior crest, located between parietals and epiotics [WV82]; parietals completely separated by supraoccipitals [$?JP97]; pterosphenoids widely separated [JP97]; extensive posttemporal fossa roofed by pterotic & parietal [JP97] (esocids only?); hyomandibula with lateral wing bracing dorsal arm of preopercular (i.e. the "opercular process" in figure at Umbra) [WV82] [JP97]; hyomandibula lateral crest short [$?JP97]; metapterygoid located posterodorsal to quadrate [JP97] (contra [WV82]: figured as dorsal at least in Umbra); prominent paired dermal ethmoid bones (proethmoids) [R74]; proethmoids elongate [JP97]; orbitosphenoid absent [JP97]; palatines fused to autopalatine [JP97]; maxilla toothless but in gape of mouth [W84] [JP97]; endopterygoid (= pterygoid) without teeth [JP97]; fused anguloarticular [JP97]; lower jaw from dentary, retroarticular, anguloarticular & coronomeckelian [WV82]; anguloarticular forms entire joint with quadrate [WV82]; articular reduced [$?JP97]; retroarticular with ligamentous connection to interopercular & epihyal [WV82]; ceratohyal, dorsal margin encased in bone [$?JP97]; long anterior pharyngeal toothplate divided into 2 sections [R74] [$JP97]; 1st pharyngobranchial conical, with "tip closed in bone" (not cartilaginous) [JP97]; 2nd pharyngobranchial toothplate absent [$?JP97]; in arches 4-5, 4th basibranchial with well-developed toothplate, 5th with none [R74] [$JP97]; 4th epibranchial primitively? with uncinate process [R74] [$?JP97]; 5th epibranchial absent [R74];
5th pharyngobranchial toothplate absent [JP97]; long, highly-muscled body; S-shaped supraneurals [W84]; no adipose fin & dorsal and anal fins located posteriorly & opposite each other [R74] [W84] [$?JP97] [N94]; no fusion of caudal centra [R74]; neural arches of preurals 2-3 subequal [$?JP97]; 0-1 ural neural arches [R74]; no specializations of caudal neural or hemal spines [R74]; single urodermal present, usually extending to first preural centrum, usually (except some individuals of Umbra and Dallia) with 1+ acuminate processes anterodorsally [R74] [JP97] [1]; "urodermal" absent [$?JP97] [3]; caudal median cartilage absent [$?JP97]; caudal scutes absent [$?JP97]; internal (intercalar) limb of posttemporal "typically" present [JP97]; cleithrum ventral (coracoid) process absent [JP97]; single postcleithrum "positionally homologous with" 3rd postcleithrum [$JP97] (but [WV82]: absent in Dallia); no mesocoracoid [N94]; posterior pubic symphysis absent [JP97]; no pelvic radials [JP97]; acellular bone present (except Novumbra?) [JP97]; no breeding tubercules [JP97]; cheek & operculum have scales [JP97]; physostomous swimbladder; no pyloric caeca [N94]; all freshwater [$JP97]; ambush predators.
Characters: maxilla toothless but included in gape [N94]; single supramaxilla [$?JP97]; median unpaired supraoccipital with posterior crest, located between parietals and epiotics [WV82]; parietals completely separated by supraoccipitals [$?JP97]; pterosphenoids widely separated [JP97]; extensive posttemporal fossa roofed by pterotic & parietal [JP97] (esocids only?); hyomandibula with lateral wing bracing dorsal arm of preopercular (i.e. the "opercular process" in figure at Umbra) [WV82] [JP97]; hyomandibula lateral crest short [$?JP97]; metapterygoid located posterodorsal to quadrate [JP97] (contra [WV82]: figured as dorsal at least in Umbra); prominent paired dermal ethmoid bones (proethmoids) [R74]; proethmoids elongate [JP97]; orbitosphenoid absent [JP97]; palatines fused to autopalatine [JP97]; maxilla toothless but in gape of mouth [W84] [JP97]; endopterygoid (= pterygoid) without teeth [JP97]; fused anguloarticular [JP97]; lower jaw from dentary, retroarticular, anguloarticular & coronomeckelian [WV82]; anguloarticular forms entire joint with quadrate [WV82]; articular reduced [$?JP97]; retroarticular with ligamentous connection to interopercular & epihyal [WV82]; ceratohyal, dorsal margin encased in bone [$?JP97]; long anterior pharyngeal toothplate divided into 2 sections [R74] [$JP97]; 1st pharyngobranchial conical, with "tip closed in bone" (not cartilaginous) [JP97]; 2nd pharyngobranchial toothplate absent [$?JP97]; in arches 4-5, 4th basibranchial with well-developed toothplate, 5th with none [R74] [$JP97]; 4th epibranchial primitively? with uncinate process [R74] [$?JP97]; 5th epibranchial absent [R74];
5th pharyngobranchial toothplate absent [JP97]; long, highly-muscled body; S-shaped supraneurals [W84]; no adipose fin & dorsal and anal fins located posteriorly & opposite each other [R74] [W84] [$?JP97] [N94]; no fusion of caudal centra [R74]; neural arches of preurals 2-3 subequal [$?JP97]; 0-1 ural neural arches [R74]; no specializations of caudal neural or hemal spines [R74]; single urodermal present, usually extending to first preural centrum, usually (except some individuals of Umbra and Dallia) with 1+ acuminate processes anterodorsally [R74] [JP97] [1]; "urodermal" absent [$?JP97] [3]; caudal median cartilage absent [$?JP97]; caudal scutes absent [$?JP97]; internal (intercalar) limb of posttemporal "typically" present [JP97]; cleithrum ventral (coracoid) process absent [JP97]; single postcleithrum "positionally homologous with" 3rd postcleithrum [$JP97] (but [WV82]: absent in Dallia); no mesocoracoid [N94]; posterior pubic symphysis absent [JP97]; no pelvic radials [JP97]; acellular bone present (except Novumbra?) [JP97]; no breeding tubercules [JP97]; cheek & operculum have scales [JP97]; physostomous swimbladder; no pyloric caeca [N94]; all freshwater [$JP97]; ambush predators.
Characters-2: The following list was compiled as characters of "Umbridae" conceived as a clade including Umbra, Novumbra, and Dallia. Under the current scheme, it ought to represent the plesiomorphic state of Esociformes.
Characters: 10-35 cm; snout short & well rounded [N94]; nasals strongly reduced or absent [WV82]; upper jaw extends to anterior margin of orbit [WV82]; maxilla with peg-like head, lacking palatine process [WV82]; frontals large, major roofing bone [WV82]; frontals taper anteriorly & underlie proethmoid [WV82]; frontal, lateral margin concave above orbit, between lateral ethmoid & ![Umbrid dorsal skulls from [WV82]](images/UmbridDorsalSkulls.gif) sphenotic [WV82]; frontals widest at sphenotic (just postorbitally), tapers & ends at supraoccipital [WV82]; parietals reduced and separated by supraoccipital [WV82]; posttemporal forked, with dorsal limb joins epiotic & ventral joined exoccipital by ligament [WV82]; posttemporal overlies supracleithrum [WV82]; symplectic large & slightly bent [WV82]; two arms of symplectic of subequal length [JP97]; preoperculum with dorsal & ventral arms meeting at right angle [WV82]; preopercular, dorsal arm, braced on hyomandibular, dorsal process, while ventral arm against ventral surface of quadrate, quadratojugal process [WV82]; quadrate large, with prominent notch in articular condyle & prominent quadratojugal process grooved ventrally for anterodorsal edge of preopercular [WV82]; occipital condyle centrum-like, formed only from basioccipital [WV82]; basioccipital M-shaped in transverse section (?) [WV82]; basioccipital without neural arch or parapophyses [WV82]; basisphenoid absent [JP97]; C1 centrum & basioccipital joined by tubular ligament [WV82]; foramen magnum bounded by basioccipital ventrally, exoccipitals laterally [WV82]; exoccipitals with ventromedial process bordering foramen magnum & broadly contacting basioccipital [WV82]; exoccipital with slender anteroventral process medial to sacculus [WV82]; exoccipital also participates in lateral wall of sacculus and contacts epiotic via cone-shaped anterodorsal process bearing large vagus foramen [WV82]; epiotic in form of 3 cones joined at apex, with bases projecting anteroventrally, posteroventrally (both contacting exoccipital) & anteromedially [WV82]; pterotic forms much of wall of the posttemporal fossa [WV82]; pterotic, dermal portion, also extends skull table laterally (PTO in figure) and bears temporal sensory canal (TSC in figure) [WV82]; intercalar absent [WV82]; prootic large, articulating with pterotic posterodorsally, hyomandibula dorsally, alisphenoid & sphenotic anterodorsally, but no contact with exoccipital [WV82]; prootic with posterodorsal & posteroventral concavities participating in posttemporal fossa and saccular recess, respectively [WV82]; prootic anterior face forms posterior wall of orbit [WV82]; elongate, waisted alisphenoid joins frontal, prootic & sphenotic [WV82]; sphenotics cone-shaped, forming
sphenotic [WV82]; frontals widest at sphenotic (just postorbitally), tapers & ends at supraoccipital [WV82]; parietals reduced and separated by supraoccipital [WV82]; posttemporal forked, with dorsal limb joins epiotic & ventral joined exoccipital by ligament [WV82]; posttemporal overlies supracleithrum [WV82]; symplectic large & slightly bent [WV82]; two arms of symplectic of subequal length [JP97]; preoperculum with dorsal & ventral arms meeting at right angle [WV82]; preopercular, dorsal arm, braced on hyomandibular, dorsal process, while ventral arm against ventral surface of quadrate, quadratojugal process [WV82]; quadrate large, with prominent notch in articular condyle & prominent quadratojugal process grooved ventrally for anterodorsal edge of preopercular [WV82]; occipital condyle centrum-like, formed only from basioccipital [WV82]; basioccipital M-shaped in transverse section (?) [WV82]; basioccipital without neural arch or parapophyses [WV82]; basisphenoid absent [JP97]; C1 centrum & basioccipital joined by tubular ligament [WV82]; foramen magnum bounded by basioccipital ventrally, exoccipitals laterally [WV82]; exoccipitals with ventromedial process bordering foramen magnum & broadly contacting basioccipital [WV82]; exoccipital with slender anteroventral process medial to sacculus [WV82]; exoccipital also participates in lateral wall of sacculus and contacts epiotic via cone-shaped anterodorsal process bearing large vagus foramen [WV82]; epiotic in form of 3 cones joined at apex, with bases projecting anteroventrally, posteroventrally (both contacting exoccipital) & anteromedially [WV82]; pterotic forms much of wall of the posttemporal fossa [WV82]; pterotic, dermal portion, also extends skull table laterally (PTO in figure) and bears temporal sensory canal (TSC in figure) [WV82]; intercalar absent [WV82]; prootic large, articulating with pterotic posterodorsally, hyomandibula dorsally, alisphenoid & sphenotic anterodorsally, but no contact with exoccipital [WV82]; prootic with posterodorsal & posteroventral concavities participating in posttemporal fossa and saccular recess, respectively [WV82]; prootic anterior face forms posterior wall of orbit [WV82]; elongate, waisted alisphenoid joins frontal, prootic & sphenotic [WV82]; sphenotics cone-shaped, forming ![Umbrid palatal skulls from [WV82]](images/UmbridPalatalSkulls.gif) posterolateral corner of orbit & part of hyomandibular articulation posteroventrally [WV82]; ethmoids present as two paired elements: proethmoids & (perichondral) lateral ethmoids [WV82]; proethmoids thin & plate-like, overlying frontals posteriorly [4] [WV82]; lateral ethmoids are flattened cones capping posterolateral corners of rostral cartilage [WV82]; mesopterygoid large, extending far medially [WV82] [5]; metapterygoids with vertical ventral plate joined to a medially inclined dorsal plate [WV82]; ectopterygoids reduced [WV82]; maxilla edentulous [WV82]; depressible teeth primitively present? (Palaeoesox) [W+92]; dentary bears teeth for about 2/3rds of length [WV82]; mandibular sensory canal relatively large compared to esocids) [W+92]; branchiostegal rays 5-8 [N94]; hyomandibula with single head articulating posterodorsally with pterotic, anterodorsally with sphenotic, & anteriorly with prootic (?) [WV82]; hyomandibular, anteroventral wing from shaft underlies posterior metapterygoid & lateral wing contacts preopercular [WV82]; interhyal short [WV82]; epihyal short & plate-like [WV82]; ceratohyal waisted and thick [WV82]; two hypohyals, both small & rounded [WV82]; single main urohyal present [WV82]; branchial arches infrapharyngobranchials (4 pairs), epibranchials (4 prs), ceratobranchials (5 prs), hypobranchials (3 prs), and basibranchials (4 prs), plus a basihyal [WV82]; basibranchial & infrapharyngobranchial of 4th arch not ossified [WV82]; 1st & 2nd basibranchials always lack toothplates [WV82] contra [R74]: toothplates on 1st & 3rd); ossified gill rakers on all except posterior of 5th arch [WV82]; body long & slender, cylindrical anteriorly & laterally compressed caudally [PB91]; 32-42 vertebrae [N94]; single dorsal fin & anal fins posteriorly placed [PB91]; neural arches fused to centra [JP97]; epineurals not fused to arches [JP97]; caudal fin rounded [N94] (not true of Novumbra); caudal skeleton with 2 urals, 4-7 hypurals, 1 pair of uroneurals and 0-2 epurals [WV82]; caudal rays 20-30, with 8-19 branched; fin rays absent [PB91]; some marked individual variation in caudal skeleton [WV82]; pelvic fins small & abdominal [PB91]; scales cycloid [PB91]; scales small, with central focus and fine concentric circuli [WV82]; lateral line faint, if present; pores in infraorbital canal 3 or less; chromosome number very variable (22-78); said to be able to survive on atmospheric oxygen alone; also highly resistant to both high & low temperatures and even absence of water; does not tolerate brackish conditions; eat insects, snails, crustaceans, and crayfishes (notably good at controlling mosquito larvae).
posterolateral corner of orbit & part of hyomandibular articulation posteroventrally [WV82]; ethmoids present as two paired elements: proethmoids & (perichondral) lateral ethmoids [WV82]; proethmoids thin & plate-like, overlying frontals posteriorly [4] [WV82]; lateral ethmoids are flattened cones capping posterolateral corners of rostral cartilage [WV82]; mesopterygoid large, extending far medially [WV82] [5]; metapterygoids with vertical ventral plate joined to a medially inclined dorsal plate [WV82]; ectopterygoids reduced [WV82]; maxilla edentulous [WV82]; depressible teeth primitively present? (Palaeoesox) [W+92]; dentary bears teeth for about 2/3rds of length [WV82]; mandibular sensory canal relatively large compared to esocids) [W+92]; branchiostegal rays 5-8 [N94]; hyomandibula with single head articulating posterodorsally with pterotic, anterodorsally with sphenotic, & anteriorly with prootic (?) [WV82]; hyomandibular, anteroventral wing from shaft underlies posterior metapterygoid & lateral wing contacts preopercular [WV82]; interhyal short [WV82]; epihyal short & plate-like [WV82]; ceratohyal waisted and thick [WV82]; two hypohyals, both small & rounded [WV82]; single main urohyal present [WV82]; branchial arches infrapharyngobranchials (4 pairs), epibranchials (4 prs), ceratobranchials (5 prs), hypobranchials (3 prs), and basibranchials (4 prs), plus a basihyal [WV82]; basibranchial & infrapharyngobranchial of 4th arch not ossified [WV82]; 1st & 2nd basibranchials always lack toothplates [WV82] contra [R74]: toothplates on 1st & 3rd); ossified gill rakers on all except posterior of 5th arch [WV82]; body long & slender, cylindrical anteriorly & laterally compressed caudally [PB91]; 32-42 vertebrae [N94]; single dorsal fin & anal fins posteriorly placed [PB91]; neural arches fused to centra [JP97]; epineurals not fused to arches [JP97]; caudal fin rounded [N94] (not true of Novumbra); caudal skeleton with 2 urals, 4-7 hypurals, 1 pair of uroneurals and 0-2 epurals [WV82]; caudal rays 20-30, with 8-19 branched; fin rays absent [PB91]; some marked individual variation in caudal skeleton [WV82]; pelvic fins small & abdominal [PB91]; scales cycloid [PB91]; scales small, with central focus and fine concentric circuli [WV82]; lateral line faint, if present; pores in infraorbital canal 3 or less; chromosome number very variable (22-78); said to be able to survive on atmospheric oxygen alone; also highly resistant to both high & low temperatures and even absence of water; does not tolerate brackish conditions; eat insects, snails, crustaceans, and crayfishes (notably good at controlling mosquito larvae).
 Notes: [1] Rosen [R74] suggests that these processes incorporate the ural neural arch of preural 1 where that arch is absent. This seems unlikely for several reasons. First, the urodermals (f/k/a "uroneurals") are derived from scales, not endochondral bone. Second, the absence of a full neural arch is associated with the presence of an additional epural, which fully accounts for the "missing" arch. [2] There is probably a third family of Esociformes, Palaeoesocidae, based on †Palaeoesox and reasonably well represented in the Russian geological record. However, they have been even less well-studied than the esocids. This family is probably more primitive than the esocids. [3] [JP97] distinguish between uroneurals and urodermals. We are uncertain what a urodermal is in their terminology. [4] The proethmoids act functionally as nasals. They are not sutured to the frontals, but closely bound by ligaments to the overlying frontals. [5] for characters of the suspensorium, refer to the image at Umbra. For the hyoid arch, see image at Novumbra.
Notes: [1] Rosen [R74] suggests that these processes incorporate the ural neural arch of preural 1 where that arch is absent. This seems unlikely for several reasons. First, the urodermals (f/k/a "uroneurals") are derived from scales, not endochondral bone. Second, the absence of a full neural arch is associated with the presence of an additional epural, which fully accounts for the "missing" arch. [2] There is probably a third family of Esociformes, Palaeoesocidae, based on †Palaeoesox and reasonably well represented in the Russian geological record. However, they have been even less well-studied than the esocids. This family is probably more primitive than the esocids. [3] [JP97] distinguish between uroneurals and urodermals. We are uncertain what a urodermal is in their terminology. [4] The proethmoids act functionally as nasals. They are not sutured to the frontals, but closely bound by ligaments to the overlying frontals. [5] for characters of the suspensorium, refer to the image at Umbra. For the hyoid arch, see image at Novumbra.
Links: Teleostei; IWR: Taxa: Esociformes; Esociformes; Discover the Fishes of All of Canada; 2orti.pdf; Gli Esociformi, Esociformes (photos); University of Alberta; Systematics of major fish groups V; Süßwasserfische Mitteleuropas; lecture15; The Virtual Aquarium of Virginia Tech--Mudminnows; Mudminnow Family, Umbridae, central mudminnow, umbra limi; Family Summary -- Fishbase; lecture15; InterNevod: СЕМЕЙСТВО УМБРОВЫЕ (UMBRIDAE) Russian); Gli Umbridi, Umbridae Italian: good image); The Distribution of Stream Fish (Dunno what these results signify, but interesting); mudminnowpage; Neue Tiere in Deutschland Steckbriefe Amerikanischer Hundsfisch ....
References: Johnson & Patterson (1997) [JP97]; Nelson 1994) [N94]; Page & Burr (1991) [PB91]; Rosen (1974) [R74]; Wilson (1984) [W84]; Wilson & Veilleux (1982) [WV82] Wilson et al. (1992) [W+92]. ATW080122
checked ATW080122.

