|
|
Permian Period |
| Paleozoic Era |
Permian: 2: Tetrapods, and Biostratigraphy |
The Permian Period: 2: Tetrapods, and Biostratigraphy
 The great diversity of near-shore and fresh water Chondrichthyans which characterized the Carboniferous began to decline during the Permian. In the oceans, xenacanth sharks dominated until the Guadalupian, when they were replaced by hybodonts. A few acanthodians also lingered into the Cisuralian. Lungfishes and coelacanths were more diverse than they are today, but all of the other Sarcopterygian fishes had already become extinct. The Permian oceans were dominated by a diverse group of spiny-finned (actinopterygian) fishes, most of which had thick, heavy scales and rather basic jaw structures if you look carefully, you can see this in the image of Palaeoniscus). Neopterygians with more derived jaw structures probably began to appear by the end of the Lopingian.
The great diversity of near-shore and fresh water Chondrichthyans which characterized the Carboniferous began to decline during the Permian. In the oceans, xenacanth sharks dominated until the Guadalupian, when they were replaced by hybodonts. A few acanthodians also lingered into the Cisuralian. Lungfishes and coelacanths were more diverse than they are today, but all of the other Sarcopterygian fishes had already become extinct. The Permian oceans were dominated by a diverse group of spiny-finned (actinopterygian) fishes, most of which had thick, heavy scales and rather basic jaw structures if you look carefully, you can see this in the image of Palaeoniscus). Neopterygians with more derived jaw structures probably began to appear by the end of the Lopingian.
The increasing aridity of the Permian not only affected plants. The tetrapods suffered as their swamps and pools shrunk and dried out. Those surviving forms included big-headed temnospondyls two to three meters in length, as well as long-snouted forms (the archegosaurs) superficially resembling small crocodiles. Many of the non-amniote reptilomorphs, such as anthracosaurs and embolomeres, continued in to the Permian.
But it was the amniotes that took over as the dominant land animals, adapted to life on land (thanks to water-retentive dry skin and the amniotic egg). Although there were a number of different types of amniotes, the largest and most diverse belonged to the Synapsida, which were ancestral to the mammals.
There were several distinct evolutionary dynasties of synapsids as the Permian progressed.
 The first, the pelycosaur dynasty, included the large finbacks of the early Permian such as Dimetrodon, Edaphosaurus, Ctenospondylus, and Secodontosaurus, all of which attained a lengths some 3 meters, as well a similar types that lacked a "sail". The large dorsal "sails" were most certainly thermoregulatory devices that would heat up the animal in the cold morning, making it more active and giving it an advantage over it's more sluggish sail-less relatives. These animals were limited to the equatorial tropics.
The first, the pelycosaur dynasty, included the large finbacks of the early Permian such as Dimetrodon, Edaphosaurus, Ctenospondylus, and Secodontosaurus, all of which attained a lengths some 3 meters, as well a similar types that lacked a "sail". The large dorsal "sails" were most certainly thermoregulatory devices that would heat up the animal in the cold morning, making it more active and giving it an advantage over it's more sluggish sail-less relatives. These animals were limited to the equatorial tropics.
 Following this was the Dinocephalian dynasty of the middle Permian (Guadalupian epoch). The Dinocephalians were among the most primitive of the therapsids or "mammal-like reptiles". Some grew to huge size (3 to 5 meters) with 50 to 80 cm long heads full of wicked teeth (the name Dinocephalian means "fearsome head"). These creatures succeeded the Pelycosaurs, being both larger in size and more metabolically active. There were several different types, the primitive anteosaurs see Anteosaurus, left) being carnivores, and the
Following this was the Dinocephalian dynasty of the middle Permian (Guadalupian epoch). The Dinocephalians were among the most primitive of the therapsids or "mammal-like reptiles". Some grew to huge size (3 to 5 meters) with 50 to 80 cm long heads full of wicked teeth (the name Dinocephalian means "fearsome head"). These creatures succeeded the Pelycosaurs, being both larger in size and more metabolically active. There were several different types, the primitive anteosaurs see Anteosaurus, left) being carnivores, and the  ox-sized tapinocephalia being herbivores.
ox-sized tapinocephalia being herbivores.
 The Dinocephalians all died out suddenly, perhaps as a result of unusual climatic factors, at the end of the Guadalupian. The Therapsids that followed them were smaller, and more mammal-like. Some may even evolved fur and the ability to control their temperatures metabolically. These included the large gorgonopsians the Permian equivalent of the "saber-toothed tiger"), the small to medium-sized Therocephalia, and the herbivorous dicynodonts. These creatures had previously lived alongside the giant Dinocephalians, but came into their own when the latter had died out.
The Dinocephalians all died out suddenly, perhaps as a result of unusual climatic factors, at the end of the Guadalupian. The Therapsids that followed them were smaller, and more mammal-like. Some may even evolved fur and the ability to control their temperatures metabolically. These included the large gorgonopsians the Permian equivalent of the "saber-toothed tiger"), the small to medium-sized Therocephalia, and the herbivorous dicynodonts. These creatures had previously lived alongside the giant Dinocephalians, but came into their own when the latter had died out.
In addition there were many types of non-syanapsid amniotes. The pareiasaurs, a group of big, armored herbivores probably related to turtles, reached enormous sizes (length up to 3 meters). Smaller, lizard-like reptiles were probably common, but they are very poorly known. These included the bolosaurs and procolophonids
-- both also turtle relatives. During the Permian, the crocodile-bird lineage (Archosauromorpha) had not yet diverged from the lizard-snake (Lepidosauromorpha) clade. The primitive reptiles were represented by lizard-like captorhinids and basal diapsids. The latter included a number of marine or amphibious forms: younginiforms, Claudiosaurus, and perhaps the earliest members of the ichthyosaur group.
Image credits: Palaeoniscus from the Essex Rock & Mineral Society. Deltavjatia from the University Museum of Zoology Cambridge.
MAK 2002. Revised ATW050604.
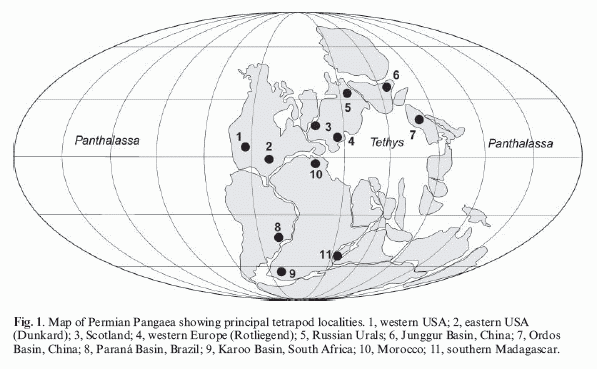
Permian tetrapod localities - from Lucas 2006 p.66.
The Permian is richly represented by fossil tetrapods, which enables us to trace the transformation of life during this important time. The main localities are in south-west United States (the famous Red Beds of texas and New Mexico), Southern Africa, the Urals region of Russia (from which the name "Permian" is derived), and more recently China, among other places, as shown in the above map. However, correlating these varying stratigraphic sequences has often proved a challenge, especially because the fauna of each tend to be highly endemic
The biostratigraphy of the Texas and New Mexico Red Bed section has been reviewed by Lucas 2006, who notes that marine biostratigraphy has shown that the youngest strata are no younger than the late Kungurian. The Kungurian in fact was a time of rapid evolutionary turn-over. The various stratigraphic stages and biozones are correlated as follows:
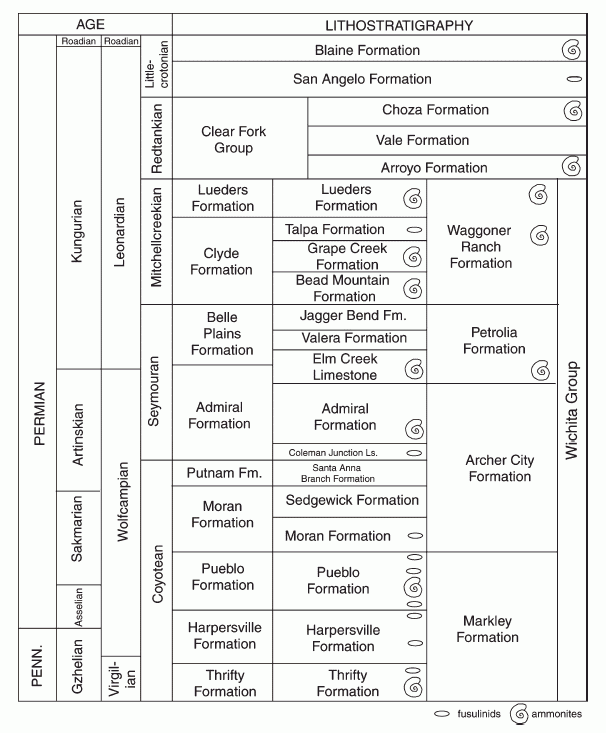
the above diagram shows Lower Permian stratigraphic section in north-central Texas showing cross-correlation of vertebrate biochronology (land vertebrate faunal zones - 4th column from the left) and marine biostratigraphy and dating (shown by fusulinids and ammonoids). Formations on right of diagram are tetrapod-bearing units. From Lucas 2006 p.71

Representative Endothiodon-Dicynodont fauna - (
Daptocephalus Zone, Lower Beaufort Series, Karoo, South Africa (late
Wuchiapingian/early
Changhsingian of south-central
Gondwana). These represent the type of animals whose remains are preserved in the Upper Permian Karoo sediments, and constitute a cross-section of the terrestrial fauna of the time.
From top left to bottom right:
Rhinesuchus (
Rhinesuchidae - a large fish-eating
Temnospondyl);
Procynosuchus (
Procynosuchidae, a small advanced
cynodont therapsid carnivore/piscivore/insectivore); an ox-sided herbivorous
Pareiasaurus (
Pareiasauridae -
anapsid herbivore); a small
diapsid Paliguana ("Paliguanidae" - diapsid/"eosuchian" insectivore); a large herbivorous
Dicynodont,
Aulacephalodon (Aulacephalodontidae therapsid herbivore); and a
Gorgonopsian therapsid,
Gorgonops (
Gorgonopsidae), which was very much the top predator of this environment
The Karroo deposits of Southern Africa are very rich in bones, especially those of therapsid synapsids, and trace the evolution of these animals over a period of some 30 million years or more. The Beaufort beds have produced a remarkable array of synapsids as well as some important archosauriform reptiles (thecodonts) and an assortment of tetrapods. Plants like Glossopteris are also well represented
Middle Permian to Early Triassic
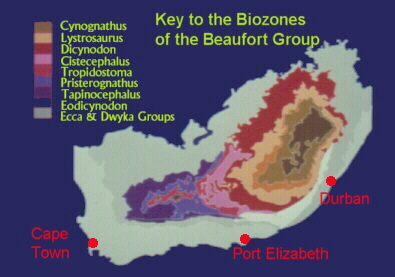
graphic from the Bernard Price Institute
Past and Present biostratigraphic divisions of the Karoo Supergroup
| Cynognathus |
Cynognathus |
Cynognathus |
Kannemeyeria |
Kannemeyeria - Diademodon |
Tetragoia |
Cynognathus |
| Kannemeyeria |
| Procolophon |
Procolophon |
Lystrosaurus |
Lystrosaurus |
Lystrosaurus - Thrinaxodon |
Lystrosaurus |
Lystrosaurus |
| Lystrosaurus |
Lystrosaurus |
| Kistecephalus |
Cistecephalus |
Daptocephalus |
Dicynodon lacertips |
Dicynodon lacertips - Whaitsia |
Dicynodon |
Dicynodon |
| Cistecephalus |
Aulacephalodon baini |
Aulacephalodon - Cistecephalus |
Cistecephalus |
Cistecephalus |
| Endothiodon |
Endothiodon |
Tropidostoma microtrema |
Tropidostoma - Endothiodon |
Tropidostoma |
| Pareiasaurus |
Tapinocephalus |
Tapinocephalus |
Pristerognathus - Diictodon |
Pristerognathus - Diictodon |
Robertia |
Pristerognathus |
| Dinocephalian |
Dinocephalian |
Tapinocephalus |
| |
|
|
|
|
|
Eodicynodon |
Table modified from - Hancox and Rubidge, 1997.
Problems of East European Stratigraphy
The region around the Ural Mountains - called the Cis-Urals by Russian paleontologists - is an important region of Permian geology and paleontology. Because of the importance of East European deposits in Permian stratigraphy (indeed, the Permian period was named by Murchison after the Perm region in eastern Russia), these became the basis for a global standard. The local geological stages here - the Asselian, Sakmarian, Artinskian, and Kungurian, were incorporated into the International stratigraphic tables, and the whole Early Permian officialy called the Cisuralian. Unfortunately, correlations with the "Upper Permian" stages are less clear, because of a paucity of index fossils (e.g. conodonts, ammonites, etc), due to these sequences being mostly terrestrial. This is unfortunate, because these rocks - rich in tetrapod fossils - trace the evolution of reptiles and amphibians throughout the middle and late Permian, in a manner like the equivalent Beaufort Series of the South African Karoo.
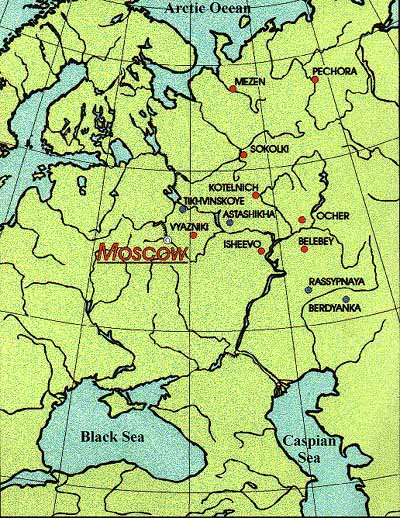
map of Permian and Triassic East European fossil localities
|
The Russian late Permian strata are traditionally divided into the Ufimian (the earliest, immediately succeeding the Kungurian), the Kazanian, and the Tartarian (the latest, corresponding to the uppermost Permian deposits). Because of the importance of East European deposits in Permian stratigraphy (indeed, the Permian period is named after the Perm region in eastern Russia), these became the basis for a global standard. Unfortunately, the exact details of where each begins and ends is unfortunately somewhat obscure.
There is some controversy regarding the end of the Tartarian. According to Kozur 1998 the Tartarian ends some considerable time before the end of the Permian, so that the uppermost Tartarian is only earliest Wuchiapingian, which seems implausible in view of the advanced tetrapod fauna. Lozovsky 1998 in contrast indicates (at least from the diagrams) the Tartarian seems to stop at the middle Changhsingian, or thereabouts. However, a more recent stratigraphic and biostratigraphic tabulations (see e.g. Benton et al 2004, Lucas 2004, Kotlyar and Pronina-Nestell 2005, Taylor et al 2009 and link) extend the Tartarian right up to the P-T (Permian-Triassic) boundary. In that table (shown below), the Tartarian includes all the substages from the Wordian upto the Wuchiapingian
The following tables are from Lucas 2004 and Kotlyar and Pronina-Nestell 2005 and present two very different correlations, especially as regards the Kazanian, and important stage for Middle Permian tetrapods in East Europe.
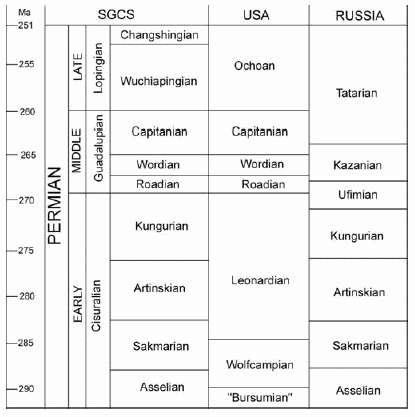
Permian marine timescales relevant to this article. The standard global chronostratigraphic scale (SGCS) is from Wardlaw (1999), as is the numerical calibration, which is tentative. Correlation of the North American and Russian scales to the SGCS is that of Glenister et al. (1992), Kozur (1995) and Kotlyar (2000).
References:
Glenister, B.F. et al. 1992. The Guadalupian. Proposed international standard for a Middle Permian Series. International Geology Review, 34: 857-888.
Kotlyar, G.V. 2000. Permian of the Russia and CIS and its interregionalcorrelation. In: Yin, H., Dickins, J. M., Shi, G. R. and Tong, J., Eds., Permian-Triassic evolution of Tethys and western circum-Pacific. Amsterdam: Elsevier, 17-35.
Kozur, H. 1995. Permian conodont zonation and its importance for the Permian stratigraphic standard scale. Geologische Palaeontologische Mitteilungen Innsbruck, 20: 165-205.
Wardlaw, B.R. 1999. Notes from the SPS chair. Permophiles, 35: 1-2.
According to the above table, the beginning of the Tatarian is around the early-middle Capitanian. However, the paleomagnetic lllawarra Reversal, an important mid-Permian marker, has been found within the Tatarian rocks of Russia (Permophiles #31 Januray 1998 pp.35-6.). Since the Illawara Reversal is located (by conodont biostratigraphic dating) just below the base of the Capitanian (i.e., the latest Wordian), the Tatarian must therefore also include all of the Capitanian and at least some of the Wordian. The following table appeared in Permophiles #46 2005.
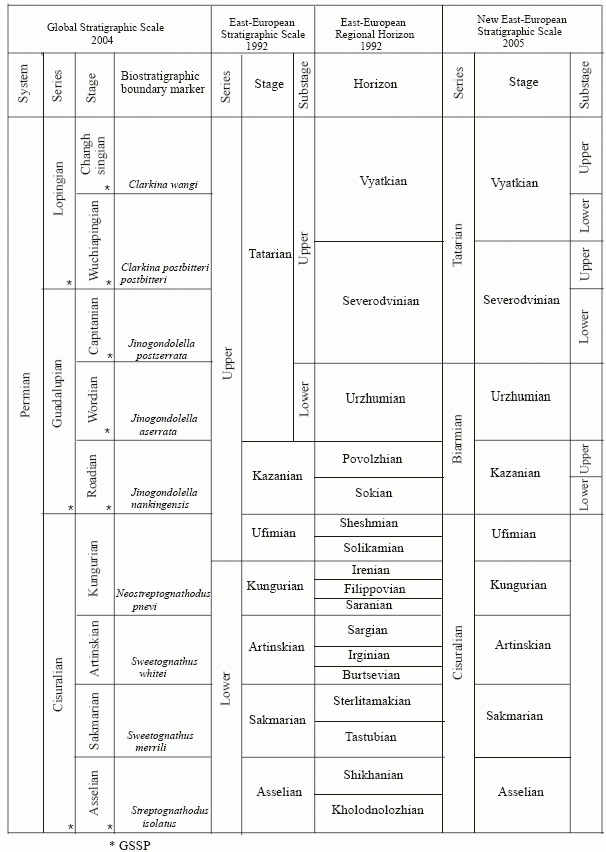
Molostovskii, 2005 argues that Upper Permian paleomagnetic zones establish that the upper Tatarian Substage of the stratotype region corresponds to the uppermost Middle–Upper Permian of the international marine scale. The Vyatka Horizon is similar in stratigraphic range to the Changhsingian, while the Severnayadvina regional stage corresponds to both the upper Capitanian Substage and the Wuchapingian Stage.
Minikh et al 2008 point to new data on asynchronous biozonal and paleomagnetic boundaries at the boundary of the Urzhumian and Severodvinian stages which show the Kiaman-Illawarra boundary occurs within the Urzhumian stage (the upper third according to Urzhumian ostracod assemblages), with Severodvinian ostracods, fishes and tetrapods only occuring far above the boundary. This implies that the upper part of the East European Urzhumian can be correlated with the upper Capitanian.
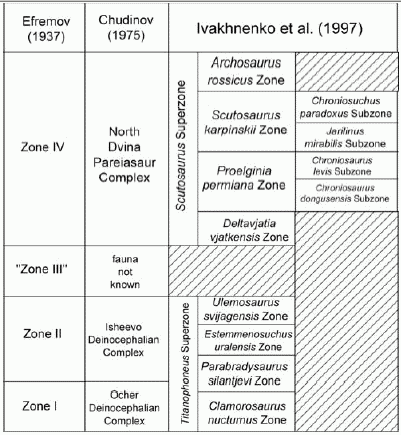
The chart on the left, from Lucas 2004, gives the various iterations of the biozonation of East European (Cis-Uralian) Tetrapods by successive Russian workers. Zone I is upper Kazanian, and hence mostly Wordian (and perhaps latest Roadian). Zone II is Urzhumian, and hence mostly Capitanian, and Zone IV is Severnayadvian and Vyatskian (Lopingian). The earlier fauna can be plotted by Dinocephalian index fossils (apart from Clamorosaurus, an endemic Euskelian temnospondyl amphibian of uncertain dating), the later faunas by Pareiasaur and reptiliomorph amphibians (Chroniosuchia). "Zone III" is non-fossilferous
The following table, based mostly on information in Olson, 1962, King, 1990, Lozovsky, 1998, and Kurkin, 2001, and Tverdokhlebov et al 2005 provides a more detailed listing of the tetrapods in East Europe. The various locality columns give the ranges for tetrapods found in those localities (see map, Permian localities in red).
| Changhsingian Age |
Tartarian |
Vyatskian |
Zone IV |
Archosaurus |
Vyazniki Assemblage Zone |
|
|
|
|
|
|
Vyazniki assemblage |
| Wuchiapingian Age |
Scutosaurus |
Sokolki Assemblage Zone |
|
|
|
|
|
Sokolki Subassemblage |
|
| Severodvinian |
Proelginia |
|
|
|
|
|
Ilinscoe Subassemblage |
|
| Deltavjatia |
|
|
|
|
Kotel'nich Subassemblage |
|
|
| Capitanian |
|
Zone III |
|
|
|
|
|
|
|
|
|
Urzhumian
(lower Tartarian) |
? ? ? ?
Zone II
|
Ulemosaurus |
? ? ? ?
Isheevian (Ulemosaurus A.Z.) |
|
|
|
? ? ? ?
Isheevo Dinocephalian Complex |
|
|
|
? ? ? ?
Upper Zone II CDC (Karglaian) |
|
|
|
|
|
| Wordian |
|
Karagalian |
|
|
|
|
|
|
| |
Bashkirian |
Lower Zone II CDC (Bashkir) |
|
? ? ? ?
Cotylosaur Complex
? ? ? ?
|
|
|
|
|
| Kazanian |
Upper |
|
|
|
|
|
|
|
|
|
Zone I
? ? ? ? |
Estemmenosuchus
Parabradysaurus |
|
Zone I CDC |
Ocher fauna
? ? ? ? |
|
|
|
|
| Roadian |
|
|
|
|
|
|
|
| Lower |
|
? ? ? ?
Clamorosaurus
? ? ? ? |
|
|
|
|
|
|
|
|
| Kungurian |
Ufimian
Kungurian |
|
|
|
|
|
|
|
|
|
|
| Artinskian |
Artinskian |
|
Zone 0 |
|
|
|
|
|
|
|
|
Here the Deltavjatia biozone can be considered either middle or late Capitanian or early Wuchiapingian, depending on how you want to interpret things.
Permian global tetrapod correlations
The following diagram by Lucas 2006 p.67 compares American (equatorial West Pangea) with African (south central Pangea) sequences.
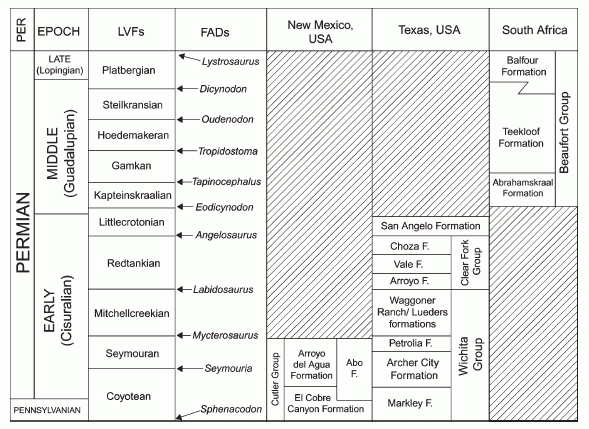
Ten land vertebrate zones are plotted, representing the transition from pelycosaur- to dinocephalian- to advanced therapsid dominated faunas. Because of the endemic nature of the Russian fauna, Lucas did not attempt a comparison. However I consider there is enough comparitive material to do so, because even if there is not a species or often no genus level match, there is a definite common pattern of faunal change, as well as climatic, paleomagnetic, and other factors.
The following useful table is from Ochev 2001 and gives Cis-Ural - Karoo correlations
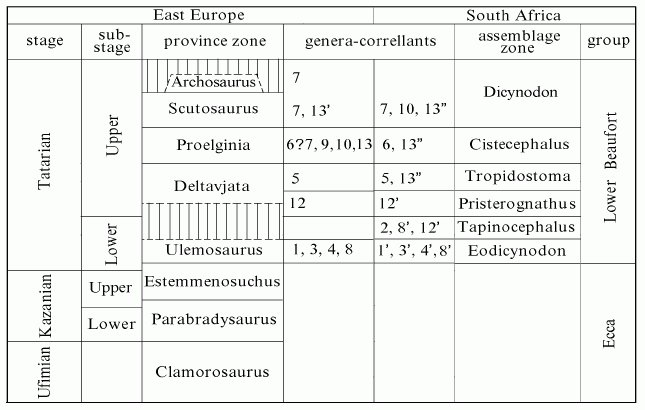
Biozone stratigraphic correlations between Eastern Europe (left) and South Africa (right). Numbers designate the genera of the families common for the regions compared. Primed numbers designate the South African genera close to the East European ones under the same numbers in their evolutionary levels. Dinocephalia: Fam. Ulemosauridae: 1 – Ulemosaurus, 1' – Tapinocaninus; Fam.Tapinocephalidae: 2 – Moschops; Fam. Anteosauridae: 3 – Titanophoneus, 3' – Australosyodon. Anomodontia: Fam. Venyukoviidae: 4 – Ulemica, 4' – Patronomodon. Fam. Dicynodontidae: 5 – Tropidostoma, 6 – Oudenodon, 7 – Dicynodon. Therocephalia: Fam. Pristerognathidae: 8 – Porosteognathus, 8' – Glanosuchus. Gorgonopia: Fam. Gorgonopidae: 9 – Sauroctonus, 9' – Scylacops. Eotheriodontia: Fam. Burnetiidae: 10 – Proburnetia, 11 – Burnetia. Pareiasaurida: Fam. Pareiasauridae: 12 – Deltavjatia, 12' – Bradysaurus, 13 – Proelginia, 13' – Scutosaurus, 13" – Pareiasaurus.
In addition, it seems implausible to place the transitional forms from the Dashankou locality in China (Xidagou Formation) in the Gamkan (equivalent to Tapinocephalus zone) Lucas 2006 does; Liu et al 2009 attribution of Roadian age is probbaly better. This would allow a further stage can be plotted between the Littlecrotonian and the Kapteinskraalian, which can (for sake of a better name) be called the "Dashankouan".
By comparing these different fossil sequences, one particular chapter in life's epic saga, life on land during the Permian, can be traced and studied in detail
CONTINUED ON NEXT PAGE
checked ATW051110, revised and updated MAK091115
text content
M. Alan Kazlev 1998-2009