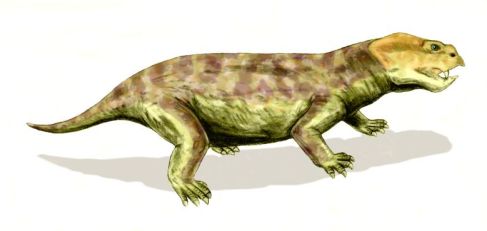
The basal dicynodont Eodicynodon. This was the first appearance of a very successful type of animal that would dominate terrestrial ecosystems right up until the end of the Cranian (late Triassic)
artwork by Arthur Weasley - Wikipedia
| Guadalupian Epoch | ||
| Paleozoic: Permian Period |
Wordian Age |
| Roadian | Cisuralian | Guadalupian | ||
| Capitanian | Lopingian | Timescale |
The Wordian stage was introduced in 1916. It is named after the Word Formation of the North American Permian Basin.
This was an important period in the history of life on Earth, as it marked the consolidation of the Roadian therapsids, and the appearance of many new forms.
Harried editor's note: I am going to be extermely lazy and quote direct from Wikipedia If you knew how many pages I have to get through here you would understand :-). Besides, why reinvent the wheel? MAK091116
The Wordian stage was introduced into scientific literature by Johan August Udden in 1916 and was named after the Word Formation of the North American Permian Basin. The Capitanian was first used as a stratigraphic subdivision of the Guadalupian in 1961, the regional timescale used for the southeastern US had the Wordian and Capitanian as subdivisions of the Guadalupian. The stage was added to the internationally used ISC timescale in 2001. The base of the Wordian stage is defined as the place in the stratigraphic record where fossils of conodont species Jinogondolella aserrata first appear. The global reference profile for this stratigraphic boundary is located at Getaway Ledge in the Guadalupe Mountains of Texas.
The top of the Wordian (the base of the Capitanian stage) is defined as the place in the stratigraphic record where the conodont species Jinogondolella postserrata first appears.
The Wordian also contains two fusulinid biozones: Afganella tereshkovae and Neoschwagerina tenuis
References: Glenister & Furnish 1961, Glenister et al 1991, Gradstein et al 2004
During the middle Wordian, , there was a reversal of the Earth's magnetic field. The Earth's magnetic field, which jhad been stable for a long time, began to flip polarities frequently. This event, known as the Kiaman-Illawarra boundary, marked the end of a long period of magnetic stability and the beginning of a period of rapid polarity shifts in the Earth's magnetic field, and may have been caused by superplume of magma deep within the Earth. This may have allowed more cosmic radiation to reach the Earth's surface, bringing about global cooling Isozakia 2009. The first magnetic flip, known as the Illawarra reversal, and is an important stratigraphic marker when dating terrestrial and marine rocks.
An aalysis by Shena et al 2009 of Roadian and Wordian occurrences of 381 brachiopod genera from 44 different geographical stations was revealed four distinct brachiopod biogeographical realms and nine provinces, and 11 brachiopod associations. The Boreal Realm in the Northern Hemisphere includes two provinces and is characterized by cold-water brachiopod associations. The Gondwanan Realm in the south also includes two provinces, the Westralian Province with biogeographical links to the Tethys. The Palaeoequatorial Realm is located mainly in the tropics and contains highly diverse and abundant brachiopod faunas. The brachiopod fauna from the Mino Belt in Japan is distinct from the other regions, and assigned to the palaeoceanic Panthalassan Realm. The major determining factor would seem to be temeprature related, with decreasing diversity from the equator to the poles. Geographic factors and oceanic currents may also have played some role.
The Wordian saw the continuation of the earlier Roadian age fauna, but with even more types of therapsids. So astonishing is this sudden evolutionary radiation that Bob Bakker refers to it as the "Kazanian Bloom". Dr Bob Bakker argues that these early therapsids were able to flourish because of their advances in metabolic development towards the mammalian condition. But in view of the fact that even advanced later therapsids like Gorgonopsids still seemed to have many ectothermic features (Freeman 1994/95) it is very unlikely that they were already partially or fully endothermic. In any case, these animals quickly radiated into an extraordinary variety of large and small terrestrial herbivores and carnivores. The Early Permian ectothermic families died out early during, or perhaps prior to, this time.
In considering Wordian faunas and communities, it seems to me that a distinction can be made between the Biarmosuchid - Estemennosuchid dominated faunas of the Latest Roadian and Early Wordian, and the more progressive dinocephalian faunas of the middle to late Wordian. These would constitute two distinct Tetrapod empires (communities)
I have assumed here that the Ocher (Ezhovo) fauna (Late Kazanian or earliest Tatarian) from which many important archaic therapsids are known, is late Roadian, although it has been also considered middle or late Wordian. (see also East European Stratigraphy Notes)

The above scene, a reconstruction of the Ocher assemblage, is probably a little later than the Belebey Community, around the Roadian/Wordian boundary. This is very similar to the Beleby fauna, except that there are larger and more diverse Estemmenosuchids, and also the presence of a number of Biarmosuchian and anteosaur carnivores, graded in size according to preditor guilds
The carnivores include the modest-sized Biarmosuchidae, relatively long-limbed lightly-built hunters of small game (a kind of therapsid dog perhaps), representing a persisting primitive lineage from which the other groups may have developed, the huge carnivorous eotitanosuchians essentially biarmosuchids grown large), the bizarre estemmenosuchids, herbivores that seem to have frequented a marshy environment, and possessing strange bony head growths, not unlike antlers, and the large brithopodids, representing another carnivorous lineage, more heavily built than the biarmosuchids. Note that apart from the estemennosuchids, which replaced the cotolyhunchines as great lumbering herbivores, all these animals were carnivores. As with the Early Permian pelycosaur-dominated fauna, this was a primitive ecosystem with a preponderance of meat-eaters over herbivores.
|
The basal dicynodont Eodicynodon. This was the first appearance of a very successful type of animal that would dominate terrestrial ecosystems right up until the end of the Cranian (late Triassic) artwork by Arthur Weasley - Wikipedia |
For a long time, the South African Beaufort Series that traces the evolution of life from the Middle Permain to the Middle Triassic was thought to only begin with the Tapinocephalus zone. In 1995 a distinct, more archaic fauna was described from the lower Abrahamskraal Formation (Rubidge 1995). This has been called the Eodicynodon Assemblage Zone. Although considered older than the Russian Zone I and II assemblages (Lucas 2004; Rubidge 1995, Lucas 2006), it would seem to me to be more advanced than the Russian Ocher and Chinese Dashankou faunas, and lence later, not earlier, as shown by the presence of primitive theriodonts (gorgonopsian, the therocephalians) and tapinophalids (specialised herbivorous dinocephalians) and the early dicynodont, the eponymous Eodicynodon itself. On the basis of its very primitive elements, Liu et al 2009 considers the Dashankou fauna Roadian, and hence older than the Eodicynodon, fauna.
 A syodont anteosaur. These medium-sized unspecialised carnivores may have had a ubiquitious distribution. artwork by Dmitry Bogdanov - Wikipedia |
This diverse fauna includes a large number of herbivores, including small primitive anomodonts (Eodicynodon, Otsheria and Patronomodon) and the huge (3 meters long) dinocephalians (Tapinocaninus). Carrnivores include the medium-sized anteosaurid dinocephalian Australosyodon (Rubidge 2004), which is very similar to Russian forms like Notosyodon and Syodon) and the more advanced and mammal-like Glanosuchus and Alopecodon; the eraliest known therocephalians. Lucas makes the Eodicynodon Assemblage Zone the characteristic assemblage for the Kapteinskraalian Land Vertebrate Faunachron (Lucas 2006)
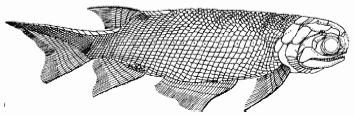 Varialepis bergi, length 34 cm;, an actinopterygian from the Bolshekinelskaya Svita of the South Cis-Urals. Order Elonichthyiformes, Family Varialepididae. From Tverdokhlebov et al 2005 p.47 |
In Russia the Bashkirian (Lower Zone II) (Olson, 1962) and Bolshekinelskaya Svita (Formation) (Tverdokhlebov et al 2005) would seem to be equivalent. The Bolshekinelskaya Formation includes Xenacanthiforme sharks, diverse Actinopterygii (right), Melosaurid temnospondyls ( Konzhukovia vetusta and Tryphosuchus sp), the Lanthanosuchid Chalcosaurus lukjanovae, and several types of Therapsida, including indeterminate Phthinosuchidae (although this is a poorly defined group in any case), carnivorous (identified only as Titanophoneus sp.) and herbivorous (Deuterosaurus jubilaei) Anteosaurids , Tapinocephalids identified as Ulemosaurus cf. gigas., but I wouldn't be superpised if they are more equivalent to the South African Tapinocaninus, the herbivorous Dinocephalian Rhopalodon(?) sp., and the Venyukoviid anomodont Ulemica efremovi (Tverdokhlebov et al 2005 p.45). The overlying (and hence more recent) Amanakskaya Svita is better represented, including Melosaurids, Archegosaurids, Seymouriamorphs, Lanthanosuchidae, Biarmosuchidae, Anteosaurids (including Syodon and the superpreditor Titanophoneus adamanteus), Tapinocephalids, Microuraniidae (Microurania minima), Venyukoviidae, and Pristerognathidae. Syodon and Porosteognathus? indicate a close connection with the Eodicynodon Assemblage Zone fauna. Differences may be due also to climatic and geograpohic factors, Russia at the tiome was very close to the equator, whereas the Karoo was in the high latitudes. Theriodonts and Dicynodonts may thus have begun as cold adapted forms, whereas the dinocephalians were probably tropical war,-weather animals, hence their greater diversity in the Cisurals.
The following diagram shows a suggested ecology of this time.
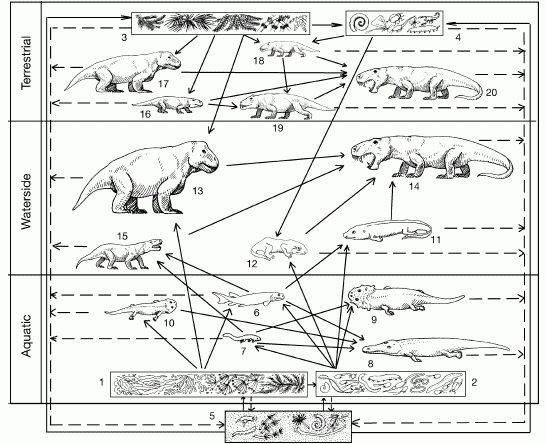
As explained by Tverdokhlebov et. al.:
Much of the Kazanian ecosystem structure survived in the early Tatarian vertebrate faunas, the Urzhumian Community, seen in the Bolshekinelskaya and Amanakskaya svitas). In the ponds and rivers, palaeonisciform, and other, fishesfed on aquatic plants and insects. But a wider community of batrachomorph amphibians, such as Konzhukovia, Uralosuchus, and Tryphosuchus, and reptiliomorphs, such as Chalcosaurus and unnamed leptorophids, preyed on the fishes, as well as on tetrapod larvae. On land, nycteroleterids and the batrachomorph Enosuchus also fed on aquatic plants and animals. The therapsid component of the fauna is much more extensive, with mediumsized herbivores such as the venyukoviid anomodont Ulemica and the dinocephalians Rhopalodon and Microurania feeding on waterside and terrestrial plants. These were preyed on by the anteosaurid dinocephalian Syodon, the therocephalian Porosteognathus and unnamed phthinosuchids. The largest herbivores are the dinocephalians Ulemosaurus and Deuterosaurus, and these were preyed on by the large anteosaurid dinocephalian Titanophoneus,, a new top-level predator.
|
Perhaps also of this same age, or a little older (Roadian) are the Various types of stem tetrapods and reptiles are well-represented in the Belebei-Mezen Cotylosaur Complex (the definition of "Cotylosaur" has since changed somewhat), which is difficult to correlate stratigraphically because of a paucity of shared faunas. It can be assumed however that numerous small lizard-like insectivorous Anapsida were an important part of the ecosystem. The Pelycosaurs may (or may not) be represented by a single femur, Phreatosaurus aenigmaticum Efremov (1954), which Efremov assigns to the family Phraetosuchidae probably an artificial group based on scrappy postcrania), but Olson, 1962 argues is really a member of the Family Caseidae, a group that is well represented in the Kungurian age.

{kind=link}
{kind=link}
{kind=link}