|
|
Reptiliomorpha |
| The Vertebrates |
Chroniosuchia |
Reptiliomorpha: Chroniosuchia
Taxa on This Page
- Bystrowianidae X
- Chroniosuchia X
- Chroniosuchidae X
The Chroniosuchia

Chroniosuchus paradoxus Vjuschkov, 1957 (family Chroniosuchidae),
life reconstruction, by Dmitry Bogdanov, 2007 (Larger image)
From Mark Mancini's blog The Theatrical Tanystropheus : Chroniosuchus:
The Chroniosuchus species were anthracosaurs which inhabited modern Russia from the latest Permian to the middle Triassic. Anthracosaurs were once considered to have belonged to the obselete “Labyrinthodontia” order. However, recent revisions concerning the cladistic relationships of early amphibians has placed the family Anthracosauridae within the superorder Reptiliomorpha and specifically inside of the order Embolomeri. It’s been suggested that anthracosaurs were closely akin to the order Seymouriamorpha due to several shared anatomcal characteristics, though many authors have questioned this and attributed their similarities to congruent evolution. In “The Age Of Dinosaurs In Russia And Mongolia”, Igor Novikov, Mikhail Shishkin and Valerii Golubev write:
“Anthracosaurs were crocodile-like piscivorous amphibians with a rather deep elongated skull from 50 to 500 mm long. During their history, they retained a resemblance to the most primitive temnospondyls in such characters as the extent of the lacrimal bone up to the naris(or septomaxilla), presence of an intertemporal (a bar of bone separating the upper and lower temporal fenestrae in reptiles) in most forms, movable basipterygoid articulation (between braincase and upper jaw), narrow interpterygoid vacuities,pterygoids with median [(as in towards the animal's midline)] contact anteriorly, a single concave occipital condoyle, occipital exposure of the opisthotic bone, and, usually, absence of retroarticular process on the lower jaw. Peculiar for anthracosaurs also is the clear demarcation boundary and loose joint between the skull roof and cheek, a condition inherited from rhipidistian fishes.”
Other features sported by anthracosaurs as noted by these authors were a lack of contact between the exoccipital and the skull roof, notably long tabular horns, comparatively narrow vomers, the presence of five digits on the forelimbs (as opposed to four in temnospondyls), and a gastrocentrous (reptile-like) spinal condition in many genera.
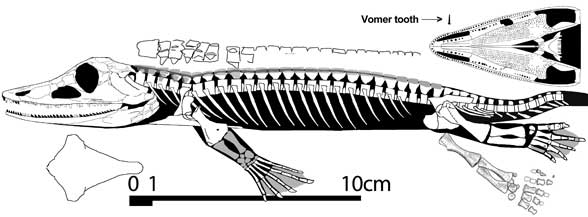
Complete specimen (THUg 3308) attributed to Chroniosaurus dongusensis by Clack and Klembara (2009), but apparently dissimilar in appearance (judging by the figures) and if so, worthy of its own generic name. ~25 cm in preserved length. - David Peters - Reptile evolution |
Chroniosuchus was, not unexpectedly, a member of the Chroniosuchia suborder, which is divided into two families: the Chroniosuchidae and the Bystrowianidae. According to Novikov et al., “They are readily distinguished from older groups by a row of dermal plates over the vertebral column, conspicuously ball-shaped [intercentrum] in most forms, fenestration (presence of holes) of the skull roof (at least in the Chroniosuchidae) and some advanced characters in the skull roof pattern.”
Unlike many earlier and contemporary amphibians, Chroniosuchids lacked a well-developed lateral line system, which indicates that these creatures likely inhabited a largely terrestrial or semiaquatic niche. The teeth of these animals were rather conical and pointed and typically displayed a slight infold at their bases. Chroniosuchids traditionally displayed additional “tusk-like teeth on their vomers, pairs or groups of small tusks on palatines and ectopterygoids.” (Novikov et al.)
However, one of the group’s most striking features was a row of sculptured osteoderm plates which ran down their backs each of which was connected (by a ligament-like structure) to its underlying vertebra via the neural arch. Adding to the intricacies accompanying the spinal cords of these beasts was the fact that their pleurocentra were amphiocoelous (concave on the anterior and posterior ends) and were, in many forms, stutured to the neural arches, and often indistinguishably so.
Copyright Mark Mancini © 2009, republished with permission
Biostratigraphy
| Period |
Regional Stage |
Zone |
Subzone |
Chroniosuchians |
Assemblage |
Age |
T
r
i
a
s
s
i
c |
|
Mastodonsaurus |
|
Synesuchus muravjevi |
|
Ladinian |
| Eryosuchus |
|
|
|
Anisian |
| Parotosuchus |
|
Axitectum georgi |
|
Olenekian |
| Vetlugian |
Benthosuchus-Wetlugasaurus |
Wetlugasaurus |
|
|
| Benthosuchus |
Dromotectum spinosum, |
|
| Tupilakosaurus |
Axitectum vjushkovi |
|
Induan |
P
e
r
m
i
a
n |
Vyatkian |
Archosaurus rossicus |
|
Uralerpeton tverdochlebovae, Bystrowiana permira |
Vyazniki |
Changhsingian |
| Scutosaurus karpinskii |
Chroniosuchus paradoxus |
C. paradoxus, C. licharevi |
Sokolki - Sokolki subassemblage |
Wuchiapingian |
| Jarilinus mirabilis |
J. mirabilis |
| Severodvinian |
Proelginia permiana |
Chroniosaurus levis |
C. levis |
Sokolki - Ilyinskoye subassemblage |
| Chroniosaurus dongusensis |
C. dongusensis |
| Deltavjatia vjatkensis |
|
Suchonica vladimiri |
Kotel'nich |
The rapid rate of chroniosuchian evolution, wide distribution and the distinctive characteristics of their armour of their remains makes them useful Eurasian index fossils. The table at the right, shows Russian late Permian biostraigraphy, consisting of four biostratigraphic zones. Three of these are represented by successive parierasaurs. Of these the middle two are subdivided according to chroniosuchid fauna, with the Proelginia permiana zone including the Chroniosaurus dongusensis and Chroniosaurus levis subzones, and the Scutosaurus karpinskii zone containing the Jarilinus mirabilis and Chroniosuchus paradoxus subzones.
During the Triassic, evolution seems to have been a lot slower, with only a few rare Bystrowianids continuing through to the end of the Middle Triassic.
References: Golubev 1998b, Golubev 1999, Shishkin et al 2000
MAK090731
Descriptions

Chroniosaurus dongusensis (Tverdokhlebova, 1972) from the Malokinelskaya Svita (Severodvinian Gorizont, upper Tatarian, Late Permian - Early Wuchiapingian age), Orenburg Region, South Cis-Urals, in lateral (a), dorsal (b), and ventral (c) views, with medial view of the lower jaw (c). From Tverdokhlebov et al. 2005 p.49 |
Chroniosuchia:
Phylogeny: Anthracosauroidea ::: * : Chroniosuchidae + Bystrowianidae. (In a recent cladistic analysis. chroniosuchids either appear as basal embolomeres, just above Silvanerpeton, with specific resemblances to embolomeres in the braincase region, or further crownward, above embolomeres but below seymouriamorphs and other stem amniotes, but here there are no unambiguous synapomorphies. Clack & Klembara 2008)
Characters: Elongated skull, but trunk short with no more than 25 praecaudal vertebrae. Dorsal armor consisting of rows of articulated scutes arranged along the midline of the back, the number of scute numbers matching the vertebrae. Scutes connected to neural spines. Lateral line canals absent. (Golubev 1998a p.283)
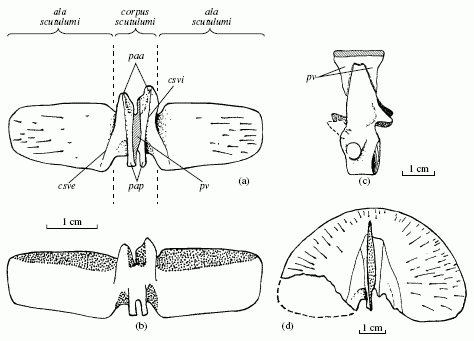
Details of Chroniosaurus armor. (a) C. dongusensis trunk scute, ventral view; (b) trunk scute showing overlap areas; (c) C. boreus trunk vertebra in left lateral view, (d) the first cervical scute of C. dongusensis ventral view. The heavy main body is the corpus scutulumi while the two lateral plates or wings are the alae scutulumi. Golubev 1998a describes the articulation and locks of the various scutes, whicha re indicated by the following abbreviations csve: crista scutulumi ventralis externa, csvi: crista scutulumi ventralis interna, paa processus articularis anterior, pap: processus articularis posterior, pv processus ventralis. From Golubev 1998a |
Armor: The dorsal osteoderms or armored scutes are roughly rectangular ins hape, except for the anterior (front) one, which is semicircular. One is reminded of Chiton (polyplacophora) molluscs. V. K. Golubev (see illustration at right) has developed a terminology for describing all the aspects of the scutes and the articulations and locks between them; those interested in such minutae are free to consult the accompanying diagram, or even better the original paper which is on line and available in English in pdf format. The articulations with the neighboring scutes are located on the ventral (under) side of the body of the scute, and is different for each of the two familuies (chroniosuchids and bystrowianiids). The dorsal (back) surface is strongly sculptured, while the ventral surface of wings is smooth and contains many small openings, which give rise to grooves and impressions of blood vessels. Golubev 1998a
Size: Skulls of Chroniosuchidae shown in Tverdokhlebov et al. (2005) are about seven to ten centimetres long, which would be only a medium-sized amphibian. However the largest specimens had skull lengths attaining 50 to 55 cm (Golubev 1998a p.283). This would imply an overall length of more than 3 meters, making these animals the dominant semi-aquatic preditors, perhaps replacing the earlier Melosaurus (link) temnospondyls (Archegosauroidea), which they would have equalled in size.
Diet: Piscivores (Tverdokhlebov et al., 2005).
Lifestyle: probably semi-terrestrial, although skull specializations that suggest a very different feeding mechanism from that of crocodiles. The interlocking scutes resemble those of some crocodilians and probably allowed a limited amount of flexing of the trunk region. (Clack & Klembara 2008)
Comments: Chroniosuchia are a group of "anthracosaur"-grade tetrapods best known from the upper Permian, though the family Bystrowianidae survived until the middle Triassic (Novikov & Shishkin, 2000). They seem to have had a superficially crocodile-like appearance with an armor of scutes, and some forms grew to quite respectable size (skull length upto 55 cm). These creatures were most common during the Late Permian, where they have stratigraphic value as index fossils, but survived well into the Triassic.
CKT070522 (the Palaeos wiki - Chroniosuchia), MAK090806
References: Golubev 1998a, Novikov et al. 2000, Tverdokhlebov et al. 2005
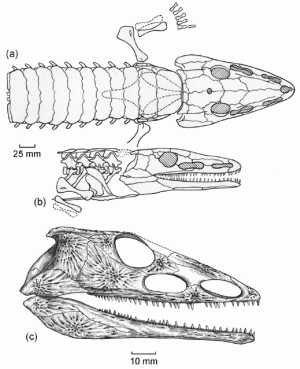
Chroniosuchus paradoxus V'yushkov, 1957 from the Kulchomovskaya and Kutulukskaya svitas Vyatkian Gorizont, upper Tatarian, Late Permian - Late Wuchiapingian age). Restorations, based on the holotype (Orenburg Region, South Cis-Urals) and skull remains, showing the anterior part of the animal, and armor plates, in (a) dorsal and (b) lateral views, and (c) lateral view of the skull. From Tverdokhlebov et al. 2005 p.50 |

Uralerpeton tverdochlebovae Golubev , 1998 Upper Tatarian Substage, Vyatkian Regional Stage (Changhsingian Age): a) and (b) the skull dorsally and laterally, reconstructed from several specimens. This was a large animal (skull lebngth 50-55 cm, one of the last of the Chroniosuchidae. From Golubev 1998a p.282 |
Chroniosuchidae: Chroniosuchus Chroniosaurus, Uralerpeton, Jarilinus, Suchonica
Phylogeny: Chroniosuchia : + Bystrowianidae + *
Characters: Large, skull length upto 50 to 55 cm. Orbits face dorsolaterally. One pair of pre-oorbital fenestrae between jugal, praefrontal, lacrymal and maxillary. Squamosal contacts postorbital. Armor wide, scute wings overlap each other posteroanteriorly. Intercentra vary from ring-shaped in young individuals to spherical in adults; the neural arch fusing with the centrum during ontogeny; see The Life of Madygen blog post.
Armor: As represented in Chroniosuchus, Jugosuchus, and Chroniosaurus, the scutes have very wide wings and cover practically the whole upper part of the trunk. The tail scutes are narrower, the last of which lack wings. A long smooth strip extends about a third of the whole wing length along the posterior dorsal edge and represents the overlap area for the next wing of the scute. This is shown in figure (b). The wings of the neighboring scutes overlap each other posteroanteriorly (the reverse to the bystrowianiid condition). The descending ventral process (processus ventralis - pv in the same figure) forms the middle part of the lock.
Reference: Golubev 1998a
Links German Wikipedia
MAK090806
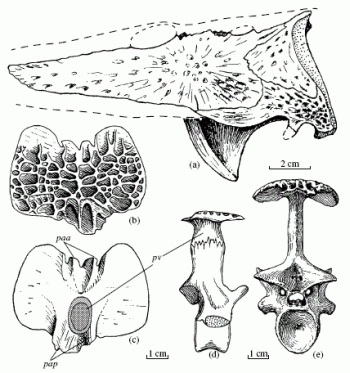
Bystrowiana permira Vjuschkov; Upper Tatarian Substage, Vyatkian Regional Stage (Changhsingian): (a) part of the skull lateral view (the anterior is to the left); (b) trunk scute dorsal view; (c) the same, ventral view; Note how short these are, in comparaison to the wide wing scutes of Chroniosaurus. (d) the trunk vertebra with fused scute from the right laterally; (e) a trunk vertebra with a fused scute posteriorly. From Golubev 1998a p.281 |
Bystrowianidae: Bystrowiana, Axitectum, Bystrowiella, Dromotectum, and Synesuchus.
Phylogeny: Chroniosuchia : + Chroniosuchidae + *
Characters: Medium sized, skull length not more than 20 cm. Orbits face dorsally. Narrow postorbital fenestra situated between skull roof and cheek. Jugal forms larger part of cheek dorsal margin. Neural arch has numerous deep pits dorsally, which sometimes turn into canals directed anteroposteriorly. Intercentra are ring-shaped or disk-shaped. Armor narrow, overlaps anteroposteriorly.
Armor: This is shown in drawings (b) and (c) (right) The osteoderms are narrow and so unlikely to be of protective value. Scute wings, if present, overlap each other anteroposteriorly (the reverse to the chroniosuchid condition), so the smooth overlap area is on the anterior edge of the dorsal surface of each scute wing. In Bystrowiana and Axitectum, the ventral processes of the neighboring scutes do not meet. Instead there are two massive posterior articulation processes. These usually fuse, forming a single articulation plate.
Comments: The only non-amniote reptilomorphs to survive the end Permian extinction event. Not very common; apart from Bystrowiana, these animals are known only by their distinctive osteoderms and a few vertebrae. Although for a long time known only from the Permotriassic of Russia, the recent discovery of osteoderms and vertebrae from the Late Ladinian (Keuper) of southern Germany show that these animals were more widespread than previously thought (Witzmann et al 2008). Bystrowiana is named after the the Russian paleontologist Alexei Petrovich Bystrow.
Reference: Golubev 1998a, Witzmann et al 2008
Links Russian Wikipedia
MAK090806
{kind=link}