 Procynosuchidae: Late Permian Cynodonts
Procynosuchidae: Late Permian Cynodonts| Cynodontia | ||
| The Vertebrates | Procynosuchidae |
| Vertebrates Home | Vertebrate | Vertebrate |
Abbreviated Dendrogram
Therapsida
│
└─Cynodontia
├─Procynosuchidae
│ ├─Nanocynodon
│ ├─Parathrinaxodon
│ └─Procynosuchus
└─┬─Galesauridae
└─Eucynodontia
├─Cynognathia
│ ├─Cynognathidae
│ └─Tritylodontidae
└─Probainognathia
├─Tritheledontidae
└─Mammaliformes |
Contents
Index |
Procynosuchidae: Late Permian CynodontsThe procynosuchids were the earliest cynodonts. They flourished during the latest Permian (Cistecephalus and Daptocephalus Zones of South Africa and Tartarian Zone IV of Russia), evolving from the same basic therapsid stock as the therocephalians. Like some of the therocephalians, the cynodonts possessed a long series of small multicusped cheek teeth and a partial secondary palate. However, the cynodonts also possessed certain unique features as well, such as a closed braincase which probably served generally to strengthen the skull and to protect the brain and middle ear cavity from pressures generated by the contraction of the increased mass of jaw muscles. The cheek teeth had a much more complex cusp pattern than that of all but a few bauriamorph therocephalians (e.g., the Bauridae). The complex cheek teeth and secondary palate show that the cynodonts were able to chew and breathe at the same time (which most reptiles cannot - they swallow their food in gulps). The food was broken down in the mouth rather than the stomach. Modifications in the jaw and the flared zygomatic arch reflect increased differentiation of jaw musculature. MAK000907. ATW030211.
 Procynosuchidae:Uralocynodon. Possibly paraphyletic stem group of Permian cynodonts.
Procynosuchidae:Uralocynodon. Possibly paraphyletic stem group of Permian cynodonts.
Late Permian of South Africa, Europe & Russia. [BS00]
Cynodontia :: Epicynodontia + * : Nanocynodon + Parathrinaxodon + Procynosuchus.
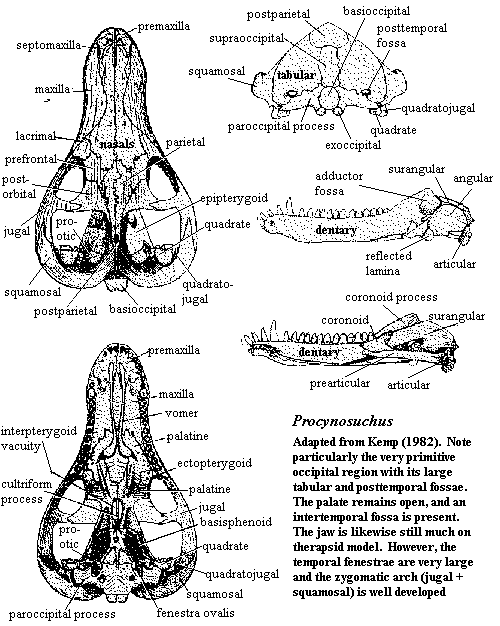
eyes generally forward-facing (possibly full stereoscopic vision); huge temporal fenestra, leaving parietal as steep sagittal crest; zygomatic arch robust and extends forward and lateral to dentary, allowing muscles to insert on lateral face of dentary [C88]; squamosal flares posteriorly in posterior wall of temporal fossa [HB86]; quadrate & quadratojugal well-exposed [BS00]; posttemporal foramen reduced [HB86]; paroccipital process expanded anteroposteriorly [HB86]; anterodorsal process of prootic extensively contacts epipterygoid [HB86]; premaxilla, palatine and maxilla expanded on palate but do not meet at midline [BS00]; probably had complete secondary palate, partially unossified [C88]; small interpterygoid vacuity present [BS00]; dentary bears coronoid process, laterally recessed for muscle insertion (i.e., a masseteric fossa) [C88]; surangular articular displaced to retroarticular process on posteroventral surface of jaw; 6 incisors & 1-2 precanines in upper jaw [BS00]; cheek teeth with multiple cusps, but linear and do not occlude [C88]; cheek teeth slight compressed laterally [BS00] cheek teeth with labial cingulum and cusps [BS00]; teeth continuously replaced [C88]; thin, mammal-like tail (so did not rely on tail for locomotion) [C88]; neither set of limbs apparently sprawled, but forelimbs may not normally have been fully extended; digits unusually long; possibly strong digging and/or swimming; insectivorous? (perhaps more diverse diet).
Links: Procynosuchus delaharpeae Broom; Procynosuchus in German).
Image: Procynosuchus, © 1999 Henssen PalaeoWerkstatt and reproduced by permission.
References: Battail & Surkov (2000) [BS00]; Carroll 1988) [C88]; Hopson & Barghusen (1986) [HB86]; Kemp (1982) [K82]; Rowe 1996) [R96]. ATW030209.
Nanocynodon: N. seductus Tatarinov 1968.
Range: Late Permian of Russia
Phylogeny: Procynosuchidae : Parathrinaxodon + Procynosuchus + *.
Characters: very small [BS00]; at least 10 postcanines [BS00]; postcanines strongly compressed transversely and sectorial [BS00]; narrow lingual cingulum with accessory cusps [BS00].
Note: may be a thrinaxodontid.
Links: RepTherapS.pdf ATW030218, MAK000907.
References: Battail & Surkov (2000) [BS00].
Parathrinaxodon: P. proops Parrington 1936. Possibly synonymous with Procynosuchus [H91].
Range: Late Permian of Africa, Kawinga Formation of the Ruhuhu Valley, Tanzania.
Phylogeny: Procynosuchidae : Nanocynodon + Procynosuchus + *.
Characters: secondary palate probably not closed [H91]; postcanine teeth identical to Procynosuchus [H91].
References: Hopson (1991) [H91]. ATW030215, MAK000907
 Procynosuchus:P. delahareae Broom. Synonyms: ?Cyrbasiodon boycei Broom 1931; Procynosuchus rubidgei Broom 1938; Paracynosuchus rubidgei Broom 1940a; Nanictosuchus melinodon Broom 1940a; Mygalesaurus platyceps Broom 1942; Aelurodraco microps Broom & Robinson 1948b; Leavachia duvenhagei Broom 1948; Galeophrys kitchingi Broom 1948; Galecranium liorhynchus Broom 1948; Suphedestes polyodon Broom 1949; Protocynodon pricei Broom 1949; Suphedocynodon gymnoternporalis Brink 1951; Leavachia microps Brink & Kitching 1951a; Leavachia gracilis Brink & Kitching 1951a; Scalopocynodon gracilis Brink 1961.
Procynosuchus:P. delahareae Broom. Synonyms: ?Cyrbasiodon boycei Broom 1931; Procynosuchus rubidgei Broom 1938; Paracynosuchus rubidgei Broom 1940a; Nanictosuchus melinodon Broom 1940a; Mygalesaurus platyceps Broom 1942; Aelurodraco microps Broom & Robinson 1948b; Leavachia duvenhagei Broom 1948; Galeophrys kitchingi Broom 1948; Galecranium liorhynchus Broom 1948; Suphedestes polyodon Broom 1949; Protocynodon pricei Broom 1949; Suphedocynodon gymnoternporalis Brink 1951; Leavachia microps Brink & Kitching 1951a; Leavachia gracilis Brink & Kitching 1951a; Scalopocynodon gracilis Brink 1961.
Range: Late Permian of Europe & South Africa, Daptocephalus Zone
Phylogeny: Procynosuchidae : Nanocynodon + Parathrinaxodon + *.
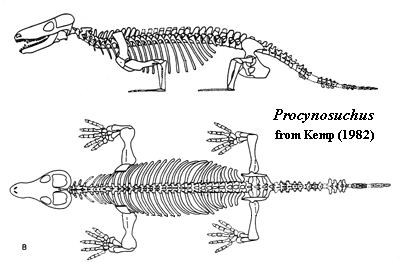
Characters: Size: 60cm long; snout broader than tall [SH98]; articular axis of quadrate somewhat anteromedially directed [K82]; quadrate and quadratojugal with simple articulations in squamosal [K82]; secondary palate very incomplete [K82]; vomers fused [K82]; tooth rows diverge posteriorly [K82]; articular surface of articular faced posterodorsally & somewhat medially [K82] [1]; 4 lower incisors [K82] [SH98]; precanine teeth on maxilla present [SH98]; anterior postcanines with a single large, slightly recurved central cusp and a labial ridge [K82]; posterior postcanines similar, but with ~5 accessory cusps on the lingual cingulum [K82]; lingual cingulum small [SH98]; buccal cingulum entirely absent [K82]; molariforms did not occlude but did shear past each other to some extent [K82]; trunk vertebrae with typical cynodont accessory zygapophyseal articulations absent [K82]; ribs without costal plates [K82]; rib heads fused into continuous synapophysis [K82]; thoracic and lumbar vertebrae somewhat differentiated, with lumbars being more massive and bearing shorter ribs [K82]; thoracic vertebrae with tight zygapophyseal articulations preventing lateral movement [K82] [2]; lumbar zygapophyses horizontal, permitting lateral undulation [K82]; posterior lumbar ribs are fused to transverse processes [K82]; may have had broad tail for swimming [K82]; limb bones & feet very flat [K82] (contra [H91] -- artifact of preservation); scapula broad & flat [K82]; coracoids large [K82]; glenoid widely open [K82] (contra [H91] -- artifact of preservation); forelimbs permanently sprawling [K82]; ulnar condyle present on humerus (not just a notch) [SH98]; ilium only slightly expanded anteriorly [K82]; hindlimbs probably capable of both sprawling & upright locomotion [K82]; insectivore? [K82]; aquatic? [K82] (contra [H91] -- supposed aquatic adaptations are all artifacts of preservation).
Note: [1] the significance of the articular angle is sketched at left. The fact that the articulation was angled implies that the musculature acting on the jaw was primarily a primitive adductor mandibulae, originating from the braincase. These muscles pull medially, and the jaw articulation is consequently angled to resist this medially- directed vector. [2] One, often repeated, speculation is that the earliest cynodonts had already acquired a diaphragm, hence the need to stiffen the thoracic region. This is also consistent with the reduction of lumbar ribs no longer needed for costal ventilation and 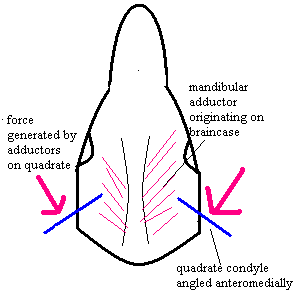 the stiffening of the thoracic rib articulations. Kemp [K82] notes an abrupt change in angle, and more gradual change in length, beginning at the 21st rib as marking the possible posterior extent of the diaphragm.
the stiffening of the thoracic rib articulations. Kemp [K82] notes an abrupt change in angle, and more gradual change in length, beginning at the 21st rib as marking the possible posterior extent of the diaphragm.
Image: right) unfortunately, it was necessary to degrade this image considerably to fit it on the page.
Links: Skelettabguß von Procynosuchus delaharpeae Broom; Procynosuchus German); Mounted skeletal reconstruction of the cynodon Procynosuchus delaharpeae Broom; Procynosuchus; Cuffey 2 - Mammal - Like Reptiles.
References: Hopson (1991) [H91]; Kemp (1982) [K82]; Sidor & Hopson (1998) [SH98]. ATW030214.
checked ATW041118