 The Occipital Face
The Occipital Face| Archosauria | ||
| The Vertebrates | Archosauriformes |
| Vertebrates Home | Vertebrate | Vertebrate |
|
Abbreviated Dendrogram
Diapsida
├─Lepidosauromorpha
└─Archosauromorpha
│
└─Archosauria (= Archisauriformes)
├─Proterosuchidae
└─┬─Erythrosuchidae
└─┬─Euparkeriidae
└─┬─Proterochampsidae
└─Crown Group Archosauria
├─Ornithodira
│ ├─Pterosauria
│ └─Dinosauromorpha
└─Crurotarsi
├─Phytosauridae
└─Rauisuchiformes
├─Rauisuchia
│ ├─Ornithosuchidae
│ └─┬─Prestosuchidae
│ └─Rauisuchidae
└─Suchia
├─Aetosauridae
└─Crocodylomorpha
|
Contents
Overview |
The Archosauriformes evolved as sprawling semi-aquatic archosaurian predators Proterosuchidae) in Pangea during the late Permian. For some fifteen million years they remained rare, undergoing very little evolutionary change. The terminal Permian catastrophe, which killed off 95% of all types of life, cleared the world of all large therapsids and allowed the proterosuchids to take center stage as the top carnivore. Within the space of five million years the proterosuchids had evolved into a wide variety of terrestrial and semi-aquatic carnivores.
Early Triassic archosauriforms like Euparkeria were the source of a further evolutionary radiation. These animals, which are historically referred to as "thecodonts", are characterized by certain shared primitive or ancestral features, such as the antorbital fenestra (an opening on each side of the skull between the eye sockets and the nostrils) and teeth in sockets. The name Thecodont is actually Latin for "socket-tooth," referring to the fact that thecodont teeth were set in sockets in the jawbones; an archosaurian characteristic that was inherited by the dinosaurs. Thecodontia therefore is an evolutionary grade of animals, rather than a clade. They represent a "grab-bag" taxon for any archosaur that wasn't a crocodilian, a pterosaur, or a dinosaur. (MAK 991003 & 030730)
The fine details of archosauriform evolution are still uncertain. In particular, Euparkeria and the proterochampsids continue to switch places as the sister group of Archosauria every few years. What is clear is that Euparkeria, Proterochampsidae, and Archosauria are a natural group and differ from other archosauriforms in a number of ways. The items on the list of synapomorphies of this unnamed group vary a bit from one study to another. We may take Paul Sereno's work as representative. Sereno (1991) includes:
1) Dorsal body armor specifically, at least one row of bony plates flanking the vertebrae down the middle of the back).
2) Interclavicle with reduced, tab-like lateral processes (as with most derived tetrapods, the structural integration of the two arms gradually decreases).
3) S-shaped femoral shaft (this one is a bit tricky, since the degree of bend in the upper leg bone is quite variable).
4) Loss of bony astragalocalcaneal canal (but it is possible that erythrosuchids have this character and proterochampsids do not -- see Chatterjee, 1982).
5) absence of ossification of distal tarsals 1 & 2
6) digit IV on the foot is markedly shorter than I.
In the 1970's and 1980's it was common to classify archosauriforms on the basis of their ankle morphology. This practice has been soundly criticized -- not least by Sereno himself. However, it is obvious from Sereno's list that there was a core of truth to the "ankle phylogenies" Sereno (1991: 2). Four of Sereno's six synapomorphies concern the structure of the hind limb. What's going on here is that both the front and hind legs are being reorganized. Conventionally, this is expressed in terms of a trend toward dinosaur-style locomotion: upright carriage with the legs held under the body and a simplified, hinge-like ankle joint. However, crocodiles do not walk in that fashion today, and basal archosaurs certainly didn't. A little later on, we will take up the matter of the archosaurian ankle in more detail. The point here is that we remain a bit uncertain exactly how the early archosauriforms, including basal archosaurs, were getting around -- just as we are unsure how the therapsids of about the same period used their legs.
Since Sereno's seminal paper, others have shown that other parts of archosauriform anatomy were also undergoing change. Gareth Dyke has gone so far as to construct archosaur phylogenies deliberately omitting ankle characters. Dyke (1998). These faithfully reproduce the main lines of descent worked out with ankle characters. From this result, Dyke argues that ankle morphology has no special significance in archosaur evolution. It may be too soon to say. Until we have a better understanding of early archosaur locomotion from a mechanical perspective, the significance of the lumps and bumps on the archosaur tarsus are difficult to evaluate.
Perhaps we should leave well enough alone with that agnostic comment. However, it may also be worth pointing out two factors which inform our uncertainty:
1. The first half of the Triassic was a period of strongly seasonal climates. In the broader context of Earth's history, this is unusual and might easily have had rather significant implications for locomotion. Survival in a strongly seasonal environment usually requires some combination of migration, torpor, and adaptability to varying substrates. Migration and broad adaptability have obvious (and different) implications for locomotion. Torpor usually requires the ability to store significant fat reserves, which has yet a third implication for locomotion. Thus, there are some a priori reasons for thinking that locomotor styles may have been under strong selective pressure -- and a type of pressure that could push in several different directions.
2. The idea that locomotor ability "improved" obscures a multitude of different adaptive strategies. Some of these strategies involve anatomical changes which are quite similar, if one simply adds up character changes, but are entirely different and incompatible in overall effect. For example, moving the legs under the body may be an adaptation for the long-distance strider who needs the speed and elevation for efficient hunting. However, the same trait may also signify adaptation for the slow-moving flat-footed walk of extremely heavy armored herbivores. The difference may show up in the foot. The strider will be digitigrade, while the plodder will be plantigrade. But a digitigrade foot may also represent a sprawling, low-slung animal adapted to sloping or uneven surfaces.
The problem with mixing similar characters which reflect incompatible locomotor designs can be seen in Sereno's synapomorphy #6: digit IV on the foot is markedly shorter than I. This wrongly lumps together animals adapted for three-toed, bipedal running with stiff-legged sprawlers [1]. There is no reason to believe that this character is homologous in all archosaurs. To the contrary, the bipedal runner is only known from the Ornithodiran lineage of archosaurs. The stiff-legged sprawler (so far as we are aware) is only known among Crurotarsi. No taxon smaller than Archosauria itself contains both types. Thus, there is no reason to suppose that this character is a true synapomorphy -- or that it even represents a single character, much less a one-time change both unique and common to the archosaur heritage.
Accordingly, we need to understand much more about the Triassic environment and early archosaur locomotion, before we can make much phylogenetic sense out of early archosaur limb characters. ATW040117.
[1] Many primitive sprawlers rotate the foot outward during stance phase. The outer digits (like digit IV) in this case end up bearing significant weight. A more stiff-legged sprawler keeps the foot pointed more or less forward, so that the weight is disproportionately carried on the inner digit (digit I).
There are about half a dozen specimens of Euparkeria, all from the Cynognathus Assemblage Zone (probably Early Anisian) in South Africa. One of these, UMCZ T692, resides at Cambridge, and its braincase has recently been described in detail by Gower & Weber (1998) [GW98]. The specimen has an interesting history. It was originally part of the private collection of DMS Watson, who did most of his field work before 1930. It was described by Rosalie Ewer in 1965, but she prepared only a portion of the braincase. In the 1970's, the braincase was extensively prepared by Arthur Cruickshank, using both acid and mechanical techniques. Shortly afterward, further mechanical preparation was performed by Alick Walker. Cruickshank is a prolific scientist. However, he was apparently interested in Euparkeria largely for comparative purposes and has never published a detailed description. Walker, we are told, chose not to publish much of his work, although he didn't mind discussing it. Benton & Gower 2002). (As a peculiar result, "A.D. Walker, personal communication" is one of the more important references in this field.)
Perhaps in consequence of this long history of preparation, the specimen seems to have literally fallen apart when Gower & Weber started to examine it in the 1990's. However, it fell apart in a most interesting and convenient way, allowing G&W an unprecedented look directly into the endocranial cavity. We will not follow G&W into these inner recesses. [2] However, we will generally follow this same pattern of deconstruction. In addition, we have the happy advantage of access to Gower's later paper, Gower (2002) [G02]. Here he lists a large number of braincase characters which may be phylogenetically significant. These characteristics will be emphasized. For those who are not braincase aficionados, it may be a good idea to review the Schematic Guide to the Mind of Bob before venturing further.
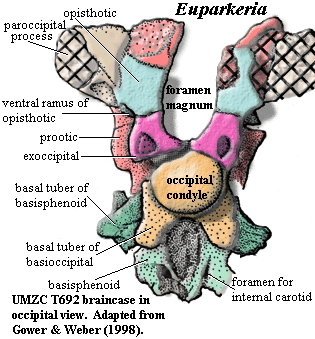
The Occipital FaceThe first figure, in occipital view, is probably close to what Ewer saw in 1965. The supraoccipital and most of the dermal bones are absent. The distal parts of the paroccipital process and the most dorsal sections of the prootic are also missing. However, the remaining parts of the occiput are relatively undistorted and undamaged.
Its a nice braincase, but not that unusual. The occipital condyle (the part that connects the skull to the back bones) is formed almost entirely by the basioccipital. The exoccipitals, which normally cooperate closely in this important enterprise, appear to be only loosely engaged. Instead they meet each other on the midline and selfishly exclude the basioccipital from the foramen magnum [G02: 5/0] [1]. The lateral edges of the exoccipitals are smoothly rounded, lacking a vertical crest [G02: 2/0]. G&W assign the large "hole" in the exoccipital to the exit of the hypoglossal nerve, the XIIth cranial nerve. That's unexpected, but not impossible. The supraoccipital is absent, but it clearly also played a major role in the foramen magnum, since the foramen is open at the top in the absence of the supraoccipital [G02: 19/0].
The opisthotic is relatively small, and is closely confined by the prootic, exoccipitals and paroccipital process. However, the ventrolateral corner is free, and a small ventral ramus descends at this point, lateral to the exoccipitals [G02: 3/0]. The ventral ramus fails to contact the prootic at any point [G02: 12/0], doubtless causing the prootic no end of worry and anxiousness.
Below the occipital condyle the basioccipital produces two substantial basal tubera. As in archosaurs, the basisphenoid attempts to confuse the issue by producing tubera of its own. In the midline, the basisphenoid also bears a large and important-looking depression which, despite appearances, is a primitive character of no known function. Of far more actual important is the location of the foramina for the internal carotid arteries. These plainly enter the braincase on the posterior face of the basisphenoid, rather than approaching from the side as in all crown group archosaurs [G02: 1/0].
 The Lateral Face
The Lateral FaceThe second figure represents the prepared lateral face of the braincase -- approximately what Cruickshank and Walker saw during the 1970's. Preparation of this area revealed that the braincase had fallen posteroventrally) away from the skull table. A fragment of laterosphenoid was attached to the parietal, but the remainder had disintegrated, as had the lateral covering of the otic capsule. Thus, in this image, we look right into the floor of the inner ear and, beyond that, into the metotic foramen between the otic capsules.
One critical feature is that there does not appear to be any well-defined (or even poorly defined) lagenar recess. This probably tells us that Euparkeria lacked perfect pitch -- or any pitch, for that matter. Mammals, crocodilians and dinosaurs independently evolved acute, pitch-sensitive hearing by elongating the lagena called the cochlea in mammals). We will omit the details, some of which may be found here. However, this form of hearing requires that a distinct perilymphatic duct receives the compression waves at the fenestra ovalis and carry them along one side of an elongate lagena in a rather specific way. In reptiles, this normally means that the inner ear must be divided into at least two compartments: (1) the scala vestibuli where the signal is received through the fenestra ovalis and in which one side (and only one side) of the lagena is open to the perilymphatic duct; and (2) a scala tympani in which the remaining energy in the compression wave carried by the perilymphatic duct is dissipated wave. There is no sign of such a system in Euparkeria and certainly no obvious recess for an enlarged lagena [G02: 9/0].
The trigeminal notch is large. A very small crista prootica is present over the facial (VII) foramen. A well-marked fossa for the palatine branch of the facial nerve trails down from the facial foramen into a large lateral depression. The anterior margin of the depression is formed by the pila antotica, and the posterior margin by a dorsal process of the basisphenoid. Anterior to this process is a "semi-lunar depression" [G02: 8/0] which G&W believe is adjacent to the fenestra ovalis. Again, this is a bit unexpected. One might expect the fenestra ovalis to be located more posterodorsally, nearer the base of the paroccipital process. In any event, the semilunar depression is a hallmark of archosauriforms, but is unknown in crown group archosaurs.
The other features of interest are mostly absence characters. The basisphenoid is not dorsoventrally tall [G02: 17/0]. The basipterygoid processes are not enlarged [G02: 18/0]. There is no horizontal ridge on the prootic below the trigeminal notch [G02: 24/1]. The basal tubera of the basisphenoid are not separated by a plate [G02: 27/1].
Euparkeria is not as typical of archosauriforms as we might want, but it does give us a basic idea of braincase geography at the base of the archosaur radiation. The otic region is longitudinally somewhat spread out by the standards of later archosaurs, and correspondingly very narrow from side to side. The walls of the capsules are not well ossified, particularly the internal (medial) face. The otic capsules of crocodilians, in particular, are highly ossified and appear to be very tightly organized -- with a form-fitting bony recess for every soft tissue element, like one of those pointless gift sets of household tools. This kind of tight spatial organization is needed for pitch-sensitive hearing because this sense depends on very precise topology. Finally, the braincase is (by croc standards) relatively unmarked by various bumps and flanges for the attachment of muscles and tendons.
If we might oversimplify -- and who can stop us? -- Euparkeria represents a sort of a plastic intermediate stage. In basal tetrapods, the braincase was as much a bony anchor for the other cranial structures as it was a convenient box for the brain. This is also true of the crocodilians, although the precise arrangements are very different. However here, at the base of the archosaurs, the braincase is relatively plastic and unconstrained. It lacks (a) an internal need to accommodate pitch-sensitive hearing through the lagena (b) an intimate relation to the dermal bones of the skull and (c) a mechanical need to anchor muscles and tendons. These are matters of degree, of course; but the relative lack of constraint may have helped set the stage for the rapid development of archosaurs in so many different directions during the critical years of the Middle Triassic. ATW031212.
[1] G&W state that the exoccipitals of Euparkeria do not meet at the midline [GW98:374]. By the time of [G02], Gower seems to have changed his mind.
[2] A variety of factors suggest that perhaps this particular specimen is too distorted to carry all the anatomical freight with which it has been loaded. We have great respect for Dr. Gower's extensive experience and observations on archosaur braincases. Perhaps because we lack that experience, the degree of crushing and distortion seems, to us, too extreme to allow a confident reconstruction of the anatomical fine points. G&W do not attempt a reconstruction, and the topographical relationships in the published figures suggest that it might be quite difficult to determine the original relationships inside the otic capsule.
Archosauriformes: Archosaurus (probably a proterosuchid).
from the Lopingian [P97]
Archosauromorpha:: Prolacertiformes + * : Proterosuchidae + Erythrosuchidae + (Euparkeriidae + (Proterochampsidae + Archosauria)).
Antorbital fenestra; orbit shaped like inverted triangle; teeth laterally compressed & serrated; widespread bipedalism; presence of 4th trochanter (Crown group only?) as attachment point for major tail muscles, the caudofemoralis group of thigh retracting muscles; astragalar facets for tibia and fibula separated by flat, non-articular surface [S91]; double row of bony scutes running along the line of the backbone; elaboration of keratin integumentary structures (e.g. feathers).
Links: link.
Reference: Jones et al. (2000), Parrish (1997) [P97], Sereno (1991) [S91]. ATW040117.
 Proterosuchidae: Archosaurus, Chasmatosaurus, Kalisuchus. Generalized primitive Archosauriforms.
Proterosuchidae: Archosaurus, Chasmatosaurus, Kalisuchus. Generalized primitive Archosauriforms.
Late Permian to Early Triassic of Russia, South Africa & Australia -- possibly South America.
Archosauriformes : Erythrosuchidae + (Euparkeriidae + (Proterochampsidae + Archosauria)) + *.
$ marked down-turn of premaxilla; supratemporal and postparietal, pineal opening, all retained; mandibular fenestra absent; shoulder girdle clearly divided as scapula & coracoid (compare Lepidosaurs); flat pelvis without thyroid fenestration; sprawling gait: head of femur terminal and directed horizontally to acetabulum.
Links: Archosaurus; Nathis Archosauriërs Dutch); Dinosaur Origins; Chasmatosaurus German); Unterordnung Proterosuchia (German -- elementary, but good); Proterosuchidae Mikko's Phylogeny); Archosaurus; Arcosauromorfa. ATW040206
 |
|
Erythrosuchus africanus. Early Triassic - length 5 meters |
Archosauriformes : Proterosuchidae + (Euparkeriidae + (Proterochampsidae + Archosauria)) + *.
Introduction: During the later part of the Early Triassic, the first thecodonts, such as Protorosuchus, gave rise both to gigantic forms Erythrosuchids) and tiny ones (euparkeriids).
Erythrosuchus africanus was five metres in overall length, with a stocky body and an enormous head a metre in length. This creature, known from the Cynognathus Zone of South Africa (similar relatives have been found in contemporary rocks in European Russia and in China) represented the culmination of early Thecodont evolution. It was obviously the largest creature of its day, being the size of a large crocodile, and must have been a fearsome predator on contemporary large herbivores, such as the abundant, bullock-sized Kanneymeriid dicynodonts.
There is a strong similarity between the Erythrosuchids of the early Triassic and the dinocephalian (primitive therapsid) anteosaurs of the Mid- Permian. In both there was a strong growth in body size, but also an even greater corresponding growth in head-size, which became fully half the length of the body excluding the tail. As with the early tetrapods, the head becomes truly enormous relative to the body. But whereas the labyrinthodonts had weak jaw and neck muscles, and had to support the weight of their great heads by the buoyancy of water, these great carnivores held their heads aloft on massive necks, and their huge teeth and tremendous jaws would have enabled them to feed easily on other animal prey.
Yet these animals, the largest predators of their time, were too heavy and clumsy to be able to actively pursue and run down prey. Like the modern crocodile or Komodo dragon, they may have been ambush-predators, lying in wait for prey behind thick vegetation, then rushing out in a short burst of speed, and quickly overwhelming the helpless victim with their tremendous jaws. (MAK 991018)
Largest terrestrial vertebrate of the Early Triassic. Dorsoventrally elongate orbit; deep maxilla beneath antorbital fossa (antorbital fossa not prominent ventral to fenestra); articulating surfaces at carpus & tarsus poorly ossified (amphibious); heavily built, to 5m; triradiate pelvis upright?).
ATW010407.
Euparkeriidae: Dorosuchus, Euparkeria. The occiput of Dorosuchus, as figured in [GS00], is strikingly similar to Euparkeria.
Early Triassic of South Africa; Middle Triassic (Anisian) of Russia.
Archosauriformes:: Proterochampsidae + Archosauria) + *.
Discussion: If the tendency to gigantism was represented by Erythrosuchus, then the opposite, tendency towards the small light agile bird-like form was expressed by its cat-sized relative Euparkeria, which at 65 centimeters (26 inches) from snout to tail-tip was only an eighth the linear dimensions. This was a small, long-legged, and agile, with a more erect stance, and capable of bipedal gait. Something similar to Euparkeria, in turn, gave rise to two evolutionary branches: the psuedosuchians and crocodiles on the one hand, and ornithodires and dinosaurs on the other. Both began as small agile forms, but the pseudosuchians quickly reverted to the standard lizard-like form, although these were lizards of great size, e.g. the phytosaurs, Aetosauridae, and Crocodylia, all of which reached 3 metres or more in length. Most interesting of all were the great rauisuchians, which although roughly lizard/crocodile-like in form had a fully erect dinosaurian and mammalian posture. MAK991018.
Small (0.5+ m); slender; facultative bipeds (?); large skull; maxillary and mandibular fenestrae present; no parietal (pineal) foramen or fossa [P93]; for braincase, see essay; foramina for internal carotids posterior to basipterygoid process [P93]; palatal teeth present [P93]; teeth laterally compressed, serrated, thecodont; vertebral intercentra present [P93]; posterior limbs 1.5x anterior, but joint surfaces etc. not specialized as biped; fourth trochanter present; mesotarsal joint (like dinosaurs) between proximal & distal tarsals (compare croc crurotarsal joint between astragalus & calcaneum) [P93]; astragalar facets for tibia and fibula separated by flat, non-articular surface [S91]; calcaneum with hemi-cylindrical facet for astragalus [P93]; calcaneal tuber convex on both sides [P93]; calcaneal tuber with some posterior deviation from lateral projection [P93]; calcaneal tuber taller than broad [P93]; calcaneal facet for fibula continuous with facet for distal tarsal IV [P93]; foot symmetrical, with elongated digit III.; dorsal row of dermal ossicles on vertebral column & tail [P93].
Introduction to Euparkeria; Lecture 8 - Tetrapods (skeletal anatomy); Triassico Italian); ???????; Archosauria: More on Morphology; Archosaurs - A new wave of Invadors; 1 Meeting of the EAVP see Borsuk-Białynicka abstract -- new Polish form); ソ。ソ・トク」トノクョセニ(Euparkeria); Euparkeria; Euparkeria capensis images of fossil); Title; Dino Web - How dinosaurs moved.
References: Gower (2002) [G02], Gower & Sennikov (2000) [GS00], Gower & Weber (1998) [GW98], Parrish (1993) [P93], Sereno 1991) [S91]. ATW030825.
Proterochampsidae: Cerritosaurus, Proterochampsa.
Middle Triassic (Ladinian) to Late Triassic (Carnian).
Archosauriformes ::: Archosauria + *.
Broad, low skull with small dorsal fenestra; no parietal (pineal) foramen or fossa [P93]; postorbital with strong, dorsally rugose, horizontal crest [S91]; foramina for internal carotids lateral to basipterygoid process [P93]; vertebral intercentra absent [S91] [P93]; primitive plate-like pelvis; calcaneal tuber with some posterior deviation from lateral projection [P93]; astragalar facets for tibia & fibula adjacent [S91]; calcaneal tuber taller than broad [P93]; calcaneal facet for fibula continuous with facet for distal tarsal IV [P93]; calcaneum with hemicylindrical facet for astragalus [P93]; dermal armor present [P93]; aquatic predators from West Gondwanaland.
Links: link treats as crocodylomorph?)
References: Parrish (1993) [P93]; Sereno (1991) [S91]. ATW040118.
checked ATW050702
Using this material. All material by ATW is public domain and may be freely used in any way (also any material jointly written by ATW and MAK). All material by MAK is licensed Creative Commons Attribution License Version 3.0, and may be freely used provided acknowedgement is given. All Wikipedia material is either Gnu Open Source or Creative Commons (see original Wikipedia page for details). Other graphics are copyright their respective owners