|
|
Synapsida |
| The Vertebrates |
Lupeosaurus,
Glaucosaurus, & Edaphosaurus |
Edaphosauridae:
Lupeosaurus, Glaucosaurus & Edaphosaurus
Taxa on This Page
- Edaphosaurus X
- Glaucosaurus X
- Lupeosaurus X

Lupeosaurus
L. kayi Romer, 1937 (type species).
Axial:
transverse processes located further above centrum than in Edaphosaurus [S89]; anterior zygapophyses relatively high above ventral margin of centra [S89]; neural spines elongated, but without tubercles [RP40] [S89]; neural spines with anticlinal bending [S89]; neural spines circular in cross-section above base [S89]; anterior cervical centra narrower than dorsal centra [S89]; cervical centra with ventral keel [S89]; cervical neural spines broadly oval & slightly constricted anteroposteriorly at their triangular base [S89]; cervical neural spines elongate [S89]; dorsal centra deeply amphicoelous, with both ends bearing reinforced rim [S89]; dorsal (but not cervical) neural spines with anterior and posterior furrows [S89]; dorsal rib with distinct heads, does not seem to flare broadly as in Edaphosaurus [S89]; sacral ribs not fused to pelvis [S89]; anterior caudal neural spines also triangular at base [S89].
Appendicular:
dorsal ramus of clavicle stout, with tongue-in-groove articulation with scapula [S89]; clavicle expanded ventrally, with thick, robust ventral plate [S89]; clavicle with broadly overlapping articulation with interclavicle [S89]; supraglenoid foramen at anterior edge of supraglenoid buttress [RP40]; scapular blade broad distally [RP40]; scapular blade curved back even more sharply than in Edaphosaurus [RP40] [S89]; scapular blade longer than in other edaphosaurs [RP40] [S89]; scapular blade with long, dorsoventrally oriented fossa on anterior edge of medial face [S89]; glenoid long [RP40]; ilium projects posteriorly more than in Edaphosaurus [RP40]; dorsal edge of pubis thickened, overhanging lateral surface [RP40]; femur short & massive [O68].
There isn't a hell of a lot to Lupeosaurus. It is known from only two described specimens, both consisting of postcranial bits and pieces. The most significant item is the absence of cross-bars on the neural spines. This left considerable doubt about the affinities of Lupeosaurus. However, the vertebrae and neural spines are otherwise entirely edaphosaurid. See details below. Unfortunately, these, and some pieces of the limb girdles, are about all that exist.
Lupeosaurus was, at a guess, in the neighborhood of 60 cm long and perhaps as much as 20 kg. The ribs suggest, but only suggest, that it was markedly skinnier than Edaphosaurus and thus not a highly adapted herbivore. On the other hand, it pushes the upper size limit for insectivores in an age when colonial insects had not yet evolved. Everything known about the limbs suggests that they were massive for an edaphosaur. The robust limbs, combined with presumed thinness, point toward a run-of-the-mill carnivore of some kind. Amphicoelous vertebral centra are a little unusual in a (relatively) powerful carnivore, but this was the primitive and usual condition for early synapsids of all types.
The skull is completely unknown.
Anteriorly. The posterior face of the centrum is more circular, and wider, than the anterior face. To judge from Sumida’s figure [S89: 1344, fig. 1E], the keel merges with a stout rim surrounding this posterior surface.
An explanation of nomenclature may be useful for the anatomically obsessive. Sumida [S89] identifies this fossa as the subcoracoscapular fossa, an attachment surface for the m. subcoracoscapularis. This muscle is the same as the m. subscapularis of mammals [LB+01], and the subcoracoscapular fossa is consequently the same as the subscapular fossa. This fossa contains a foramen which emerges on the outside (lateral face) as the supraglenoid foramen [R56].
Glaucosaurus
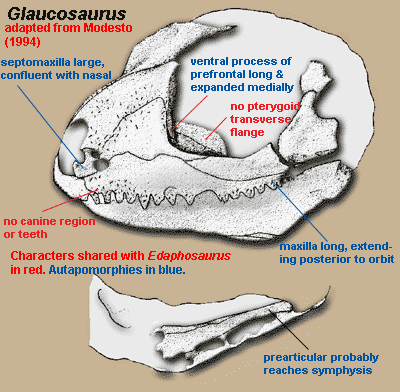
Glaucosaurus: Williston, 1915. Type: G. megalops Williston, 1915. [M94]
Range: Middle Cisuralian [L06] of Texas
Phylogeny: Edaphosauridae ::: Edaphosaurus + *.
Introduction: Glaucosaurus is known only from its holotype, a partial skull and jaw. Almost all of the sutures have been obliterated. Nevertheless, there is broad agreement that Glaucosaurus is not only an edaphosaurid, but a close relative of Edaphosaurus itself. Before getting too complacent about this phylogenetic placement, we might recall that the universe of scholars who have felt moved to exercise themselves on this topic is not large. Fortunately, that universe includes Prof. Sean Modesto of Cape Breton Univ., Nova Scotia. Prof. Modesto is merely an excellent describer of things. He also has the much rarer ability to couple description and illustration in such a manner that the reader gets a fair idea of the limitations, uncertainties and judgment calls involved.
As Modesto [M94] notes, Edaphosauridae is not marked with any cranial synapomorphies. That is, edaphosaurids don't have any unique skull features; and all we have for Glaucosaurus is a partial skull. However, Glaucosaurus seems to have two of the unique features of the clade Edaphosauridae + Sphenacodontia: (1) a lateral projection (lappet) of the frontal usually reaching the orbit and (2) ventral border of skull emarginated. Modesto asserts (credibly) that there is some suggestion that three others are also present, although it is not possible to be certain: (3) small quadratojugal which does not participate in the subtemporal bar, (4) jaw articulation below the tooth row, and (5) dentary with pronounced coronoid eminence. See image below.
All of the known sphenacodonts are carnivores except for certain therapsids such as us). Glaucosaurus is plainly not a therapsid, e.g. because the lacrimal reaches the naris, the septomaxilla is large, there are no incisors, etc. And it is just as plainly not a carnivore, since it lacks cutting edges on the teeth or canine-like teeth. So, it is very likely to be an edaphosaur. Assuming that this is the case, it is very close to Edaphosaurus, because only Glaucosaurus and Edaphosaurus completely lack both canine teeth and a canine buttress, lack the transverse flange of the pterygoid, and have prefrontal with a ventral (descending) process which is expanded toward the middle of the skull, forming an anterior housing for the eyeball [M94]. However, Glaucosaurus differs from any of the known sorts of Edaphosaurus in have an incredibly long maxilla, and in the equally extreme length of the prefrontal 's ventral process. ATW081114.
Characters:
Rostral skull: antorbital region tall [M94]; septomaxilla robust, exposed in lateral view [M94]; septomaxilla confluent with surface of nasal [M94$]; caniniform region absent [M94]; maxilla slightly convex in lateral view [M94]; maxilla extends posteriorly to level of postorbital bar [M94$].
Skull table: prefrontal ventral process expanded transversely, with extensive lacrimal contact [M94].

Lateral skull: jugal with relatively little participation in ventral skull margin [M94]; jugal lateral surface "lightly sculpted with small, circular dimples" [M94]; subtemporal bar deeper (i.e. taller) than suborbital bar [M94]; subtemporal bar emarginated [M94].
Palate: palatine with prominent dorsal process [M94]; transverse flange of pterygoid absent [M94] pterygoid dorsal process low [M94] [1].
Mandible: dentary, anterior relatively tall [M94]; splenial exposed laterally only at symphysis, underlying anterior end of angular [M94]; splenial ascends about 50% of medial jaw [M94]; inframeckelian foramen either posterior or absent [M94]; prearticular very long, extending to jaw symphysis [M94$]; coronoids toothless [M94] [2].
Dentition: 3 premaxillary teeth [M94$]; premaxillary and maxillary teeth of equal size [M94]; canines absent [M94]; 15 teeth on maxilla [M94]; dentary teeth may be smaller than maxillary [M94]; anterior maxillary teeth laterally compressed, slightly recurved, sharply tipped [M94]; posterior maxillary teeth peg-like [M94] palatal process of pterygoid toothless [M94].
Notes:
[1] Modesto [M94] states that this condition is the same in Edaphosaurus. We are not quite sure whether Modesto is referring to the dorsal ridge on the anterior ramus or (more likely) the ascending basal part of the quadrate ramus.
[2] Modesto [M94] is uncertain whether the observed structures are in fact coronoids. He notes at a number of points that the specimen has only moderately good preservation and has been seriously compromised by over-preparation. We are sympathetic, having cross-examined many witnesses with the same problems.
References: Lucas (2006) [L06]; Modesto (1994) [M94].
Edaphosaurus

Edaphosaurus: = Brachycnemius, Naosaurus)
Range: Cisuralian of central, southwestern, and eastern U.S. The range of Edaphosaurus may extend back to the latest Carboniferous [L06].
Phylogeny: Edaphosauridae ::: Glaucosaurus + *.
Introduction: Edaphosaurus possesses a combination of features that clearly indicate that this animal was a heavy, slow herbivore [RP40].
The skull of Edaphosaurus was remarkably small as compared with the size of the body. It is also short and rather shallow, as contrasted with the elongated skull of the sphenacodonts. Also unlike sphenacodonts and ophaicodonts, the teeth in the edaphosaurs are quite uniform, making an unbroken series around margins of the jaws. In addition to the marginal teeth, there is a massive array of closely packed teeth that ground against a similar set of teeth on the medial surface of the mandible. Clearly these extensive clusters of teeth added to the efficiency in grinding up plant material.
In keeping with the tiny head the cervical (neck) vertebrae are reduced in length, while the dorsal (back) vertebrae are massive, the tail is deep, the limbs are short and robust and the ribs form a wide ribcage. Like most herbivores, Edaphosaurus would have had a capacious gut and symbiotic bacteria to aid in the breakdown of cellulose and other indigestible plant material
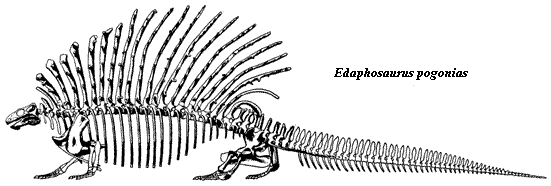
Edaphosaurus was characterized by an elongation of the vertebral spines, but the spines are heavier than they were in other forms like Echinerpeton, Lupeosaurus, and the sphenacodontids like Dimetrodon. Moreover, they were ornamented with numerous short lateral projections, tubercles, or crossbars arranged irregularly along their length, rather like the yardarms on the mast of an old sailing ship. The benefits of such an adaptation are unclear, but inasmuch as Edaphosaurus was a successful and wide-ranging animal they surely served some purpose. One possibility is that they may have served as protection against predators, by strengthening the neural spines. The front and rear spines are tilted forwards and backwards respectively, perhaps provided protection in the neck and thigh region, respectively.
Nine species of Edaphosaurus have been described, ranging in size from small (only about half a meter or less) to very large, bulky animals over three meters long). The largest species, Edaphosaurus cruciger and Edaphosaurus pogonias (see above) have modified their cervical and anterior thoracic spines into massive club-like processes. It is quite likely that Edaphosaurus evolved from a small insectivorous form like Ianthasaurus. MAK
“three consecutively occurring species, E. boanerges, E. cruciger, and E. pogonias, from Lower Permian [Cisuralian] strata of north-central Texas,” [RB86] Discussion of herbivory and jaw mechanics [M95]. Propalinal also supposed by [RP40], likewise based on having articular surface >> quadrate condyles. See also jaw of Biarmosuchus in [S03] with clear divergence between pre- and post-canine teeth.
Characters: head quite small and deep [RP40] [RB86] [M95]; skull short [M95$].
Rostral skull: snout slender [RP40]; premaxilla with dorsal process overlain by, and resting on shelf of nasal [M95]; nares small [RP40]; septomaxilla large, clearly visible in lateral view, with large medially-directed shelf probably contacting nasal septum [M95]; nasal ~75% length of frontal [M95]; nasals with strong medial suture [M95]; maxilla slightly convex in lateral view [RP40] maxilla with “lip” over teeth 10-15 [M95] [3]; lacrimal thin, underlies maxilla anteriorly, but thick, overlies maxilla posteriorly [M95]; lacrimal with strong orbital rim and strong contact with prefrontal ventral process [M95].
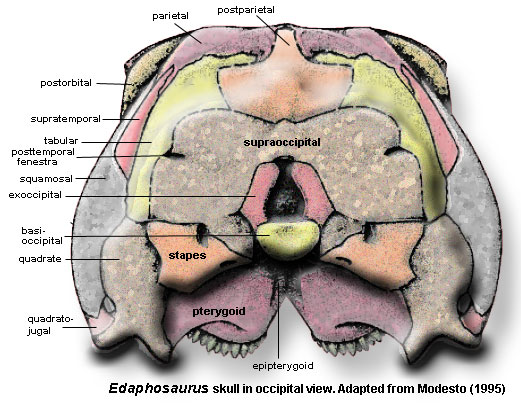
Skull table: large, well-developed supraorbital shelf formed by prefrontal (contra [RP40]), lateral lappet of frontal and postfrontal [RB86] [M95$]; prefrontal ventral process expanded transversely, with extensive lacrimal contact [M94] [M95]; prefrontal ventral process continued as internal ridge on posterodorsal process [M95]; frontal anterior process reduced [M95]; frontal lateral lappet very slender [M95]; frontal overlies parietal with serrated suture [M95]; ventral ridge from frontal-parietal suture continuous with prefrontal internal ridge [M95] [2]; postfrontal with square body forming large portion of supraorbital awning [M95] postfrontal with broad, blunt posterior process not reaching temporal fenestra [M95]; parietal narrow [RP40]; parietal with free, concave lateral margin bordering temporal fenestra [M95$]; parietal with 2 pairs of parasagittal ridges on ventral surface [M95]; parietal outer parasagittal ridge continuous with frontal ventral ridge [M95] [2]; parietal outer parasagittal ridges continuous with border of parietal foramen, then passing posteriorly to posterior flanges overlying postparietal [M95]; parietal foramen “not greatly enlarged” [RP40].
Lateral skull: jugal narrow [M95]; jugal with short anterior process meeting posterior ends of maxilla and lacrimal below orbit [RP40]; jugal with short palatal flange on anterior process [M95]; jugal with strong, extensive suture with squamosal [M95]; postorbital ventral process extending to about mid-orbit, overlapping jugal to form postorbital bar [M95]; postorbital posterior process short, not reaching squamosal or extending beyond level of parietal foramen [M95]; ventral margin of skull deeply emarginated postorbitally, with temporal bar displaced dorsally [MR95]; temporal fenestra extended (wider than tall) [M95]; temporal fenestra also tall, narrowing postorbital skull table [RP40]; squamosal with horizontal anterior process meeting jugal [M95]; ventral margin of skull (temporal bar) flares widely below temporal fenestra [RP40]; squamosal with short, rectangular occipital flange anchoring supratemporal and paroccipital process [M95]; quadratojugal small, not participating in temporal bar [M95] (contra [RP40]).
Occiput: skull table curves smoothly onto occipital surface [RP40]; parietal with occipital flange contacting tabular & postparietal [M95] postparietal with anteroventral processes fitting in slots of supraoccipital [M95]; postparietal with anterior wedge separating parietals posteriorly [RP40]; supratemporal slim, primarily occipital element [M95] [4]; tabular articulates with slot in posterior parietal & sutured to supraoccipital and paroccipital process [M95]; paroccipital process wide, extending posterolaterally & slightly ventrally, terminating with blunt, downturned tip [M95]; paroccipital process dorsal margin suturing to squamosal [RP40] [8]; exoccipitals and basioccipital fused [M95]; basioccipital forms most of semicircular occipital condyle [M95]; supraoccipital fused to (fused) otic capsule [M95]; supraoccipital with lateral processes, but poorly developed dorsal process [RP40]; posttemporal fenestra low [RP40].
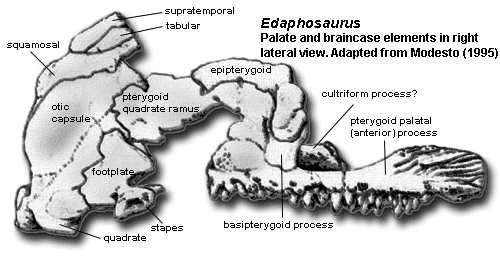
Braincase: otic capsule fused [M95]; stapes with strong dorsal process above footplate [M95]; stapedial foramen large [M95]; stapes with head small, neck distinct, & dorsal process broad [RP40]; prootics form thin, posteriorly-arching dorsum sellae [M95]; basisphenoid and parasphenoid fused [M95]; basipterygoid processes with “hour glass-shaped” double articulations [M95]; sphenethmoid (partially?) ossified, forming trough open dorsally [M95].
Palate: palatal surface of vomer with denticles posteriorly [M95]; choanae shorter and wider than in sphenacodonts [M95]; large tooth plate formed by palatine, pterygoid and ectopterygoid [M95] [5]; tooth plate slightly concave, tilted ventrolaterally [M95]; dorsal process briefly contacts lacrimal and prefrontal, forming lateral wall of (large!) lateral orbitonasal foramen [M95]; ectopterygoid small, contacts jugal briefly [M95]; pterygoid with low, deeply scarred dorsal ridge on anterior process [M95]; pterygoid transverse flange absent, modified to form semicircular posterior border of tooth plate [RP40] [M94]; pterygoid quadrate ramus thin, but with strongly elaborated proximal region rising vertically from body and wrapping around base of epipterygoid [RP40] [M95]; epipterygoid body overlies pterygoid quadrate ramus [M95]; epipterygoid with ventromedially-directed basal process forming screw-shaped receptacle for basipterygoid process of basisphenoid (i.e. the basal articulation) [M95]; epipterygoid columella (vertical process) slender with slight posterior arch [M95]; jaw suspension offset ventrally [RP40] [M95$]; quadrate condyles arranged parasagitally, separated by deep notch [M95$].
Lower jaw: lower jaw contracted and tall [RB86] [M95]; dentary and splenial with symphysial surfaces [M95]; dentary not as tall as in sphenacodonts [RP40]; dentary twisted posteriorly so that tooth row faces medially [M95]; splenial likewise twisted through almost right angle [M95]; dentary with strong suture overlapping angular and surangular [M95]; dentary forming lateral part of coronoid eminence [M95]; tooth plates formed by coronoids & prearticular [RB86]; posterior coronoid dorsal surface deeply scarred for adductor muscles [M95]; prearticular forms floor of adductor fossa [M95]; prearticular overlies medial shelf of angular, then twists ventrally to cover pterygoideus process of articular [M95]; angular large, with deep, thickened vertical keel [M95]; posterior of angular keel roughened [M95] [3]; articular surfaces of articular 50% longer than quadrate condyles [M95] [6].
Teeth: maxillary and premaxillary teeth of equal size [M94]; caniniform region absent [M94]; 21-24 isodont marginal teeth on each jaw ramus [RP40]; marginal teeth slightly bulbous [M95$]; marginal teeth small, isodont, slightly recurved, with fine, serrated tips [RB86] [M95]; cutting edges of cheek teeth inclined obliquely with respect to axis of tooth row. [M95]; marginal tooth implantation subthecodont [M95]; maxillary teeth become increasingly laterally directed posteriorly [M95$]; densely packed teeth cover tooth plates [RB86]; dentary teeth become increasingly medially directed posteriorly [M95$] [1].
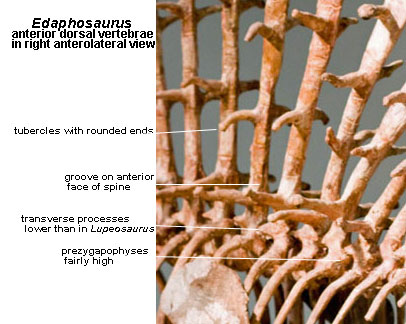 Axial: intercentra not ossified except in some posterior dorsals [RP40]; anterior zygapophyses relatively high above ventral margin of centra [S89]; neural arches not excavated [M95$]; transverse processes close to centra compared to Lupeosaurus [S89]; sail of greatly extended neural spines present [RP40]; multiple lateral tubercles, when present, usually project horizontally [M95]; lateral tubercles with rounded, unfinished tip (skin-covered), becoming smaller and less regularly paired distally [RP40]; tubercles up to 4 cm long, depending on species [H99]; neural spines extended axially at base, tapering to circular and acquiring anterior and posterior grooves (blood vessels) around first pair of tubercles [RP40]; neural spines with longitudinal grooves & ridges of variable length, originating proximally [MR90a]; anticlinal bending of neural spines, with most posterior actually curved downward distally [RP40] [9] [RB86]; 24-29 presacrals [RP40]; cervical centra short [M95$]; cervical & posterior dorsal spines slimmer [RP40]; dorsal centra longer than cervicals, relatively longer than in ophiacodonts [RP40] [RB86]; dorsal centra without ventral ridge, more or less circular in cross-section [RP40]; dorsal arches with long transverse processes [RP40] [M95$]; dorsal ribs dichocephalous [RP40]; dorsal ribs strongly curved [M95$]; dorsal rib tubercula greatly reduced [M95$]; last ~7 dorsal centra with raised tubercle for rib capitulum [RP40]; neural spines of sacrals & anterior caudals tall and pointed, with longitudinal ridges running along lateral surfaces [M95$]; 2-3 sacrals [RP40]; sacral and caudal neural spines with rugose tips [M95$]; sacral ribs not fused to pelvis [RP40]; tail probably long [RP40]; caudal neural spines tall and pointed, with tips expanded sagittally [M95$].
Axial: intercentra not ossified except in some posterior dorsals [RP40]; anterior zygapophyses relatively high above ventral margin of centra [S89]; neural arches not excavated [M95$]; transverse processes close to centra compared to Lupeosaurus [S89]; sail of greatly extended neural spines present [RP40]; multiple lateral tubercles, when present, usually project horizontally [M95]; lateral tubercles with rounded, unfinished tip (skin-covered), becoming smaller and less regularly paired distally [RP40]; tubercles up to 4 cm long, depending on species [H99]; neural spines extended axially at base, tapering to circular and acquiring anterior and posterior grooves (blood vessels) around first pair of tubercles [RP40]; neural spines with longitudinal grooves & ridges of variable length, originating proximally [MR90a]; anticlinal bending of neural spines, with most posterior actually curved downward distally [RP40] [9] [RB86]; 24-29 presacrals [RP40]; cervical centra short [M95$]; cervical & posterior dorsal spines slimmer [RP40]; dorsal centra longer than cervicals, relatively longer than in ophiacodonts [RP40] [RB86]; dorsal centra without ventral ridge, more or less circular in cross-section [RP40]; dorsal arches with long transverse processes [RP40] [M95$]; dorsal ribs dichocephalous [RP40]; dorsal ribs strongly curved [M95$]; dorsal rib tubercula greatly reduced [M95$]; last ~7 dorsal centra with raised tubercle for rib capitulum [RP40]; neural spines of sacrals & anterior caudals tall and pointed, with longitudinal ridges running along lateral surfaces [M95$]; 2-3 sacrals [RP40]; sacral and caudal neural spines with rugose tips [M95$]; sacral ribs not fused to pelvis [RP40]; tail probably long [RP40]; caudal neural spines tall and pointed, with tips expanded sagittally [M95$].
Appendicular: supraglenoid foramen above supraglenoid buttress [RP40]; scapular blade curves sharply back and is narrow distally [RP40]; cleithrum present [RP40]; clavicle broadly expanded [RP40]; “[t]he proximal limb segments are long, the distal segments short” [RP40]; ectepicondylar foramen present [RP40]; ilium very tall [RP40]; iliac blade expanded slightly [RP40]; pubis relatively short [RP40]; dorsal edge of pubis not thickened & does not overhang lateral surface [RP40]; femur short & massive [O68]; tibia head very broad [RP40]; tibia proximal lateral margin developed into sharp ridge [RP40]; tibia distal head twisted laterally “to an unusual degree” [RP40] carpals & tarsals never found in association [RP40].
Other: a 2.5 m E. boanerges estimated to be about 50% tail and a total mass of 93 kg [H99]; sail was <4% of mass but 40% of area [H99].
Notes:
[1] That is, in the back of the jaw, the teeth are not simply oriented up or down. The lower teeth tend to point inward and the upper teeth outward, with the angle increasing distally. Wear is present on the inner surfaces of the outwardly bent maxillary teeth [M95]. No one seems to know how this would actually work, especially in combination with the tooth plates. Note that the edaphosaur jaw was very deep, massive, and had a long, double symphysis (the left and right jaw were stapled together in two places). It would have virtually no ability to flex laterally. Wear on the inner surfaces of outwardly-bent maxillary teeth therefore supports Modesto’s [M95] argument that Edaphosaurus was capable of propalinal (forward and back) movement of the lower jaw. However, since at least some of the marginal teeth were not fused to the jaw (ankylosed) [M95: 221, fig. 9], the posterior teeth themselves would have flexed slightly at right angles to the jaw line.
[2] Thus, there is a continuous reinforcing arc from the orbital margin of the lacrimal, through the internal ridge of the prefrontal and onto the frontal (along the frontal-postfrontal suture?), and merging with the lateral parasagittal under the parietal. Modesto [M95: 220] states that, at least posteriorly, the ridge “presumably marks the former contact of the orbital-plate cartilage.”
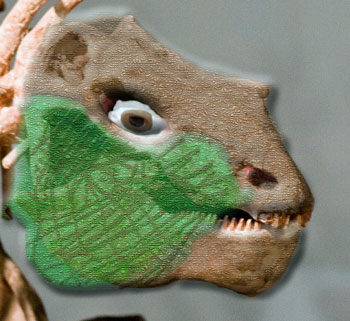
[3] One possible interpretation is attachment for a fleshy cheek, a common – in fact almost universal – feature of large herbivores. Modesto [M95] notes an attachment for an unusual muscle on the posterior third of the vertical keel of the angular. He believes that this muscle was posteriorly-directed and related to propalinal jaw movements (but where could it insert?). However, an anterior origin on the maxillary lip would have made an good, if novel, arrangement for a muscular cheek. The structure of the tooth plates almost preclude a large tongue. Consequently, it is difficult to see how serious chewing could be coordinated without strong cheeks.
[4] Contra [RP40]. Note that the supratemporal is not above the temporal region. Indeed, it does not contact the temporal fenestra at any point (being separated by the squamosal), and is barely even exposed on the temporal surface of the skull. This is an awkward, but necessary incident to the preference for homology over etymology.
[5] “tooth plates present” is given as a synapomorphy by Modesto [M95$].
[6] Romer & Price [RP40: 371] note “A further point of similarity between sphenacodontids and edaphosaurs is that the articular surface of the articular consists of two concave areas which are both elongated antero-posteriorly; [in Edaphosaurus] the inner area is much the shorter. In sphenacodontids this was associated with a corresponding elongation of the articular surfaces on the quadrate. Here, however, we have noted this was not so. Apparently the elongation in edaphosaurs was an adaptation permitting the antero-posterior movement of the lower jaw, a useful function in a herbivore.” But, if that were the case, why would the inner articulation be “much the shorter”?
[7] Romer & Price [RP40] report the type species as being E. cruciger. This seems to be incorrect. See [C06] and the discussion at [RP40: 379].
[8] This part of the skull does not seem to be well preserved in any specimen. The interpretation given by [RP40] seems reasonable, but [M95] is notably silent on the exact relations of squamosal, paroccipital process, and stapes.
[9] Romer & Price [RP40] note that the posterior dorsal spines tend to “force their way downward into the area normally occupied by the short spines of the sacrals and proximate caudals” causing the latter to be abnormally formed, twisted or even split. They emphasize that this was not pathological, since it is seen in every reasonably preserved specimen.
References: Case (1906) [C06]; Hurlburt (1999) [H99]; Lucas (2006) [L06]; Modesto (1994) [M94]; Modesto (1995) [M95]; Modesto & Reisz (1990) [MR90]; Modesto & Reisz (1990a) [MR90a]; Olson (1968) [O68]; Reisz & Berman (1986) [RB86]; Romer & Price (1940) [RP40]; Sidor (2003) [S03]; Sumida (1989) [S89].