
Life reconstruction of the Early Jurassic necked plesiosaur Plesiosaurus dolichodeirus in underwater flight.
Drawing by Dr Adam Stuart Smith (Plesiosauria com), copied from Wikipedia, Creative Commons Attribution Share-Alike
| Sauropterygia | ||
| The Vertebrates | More on Plesiosauria |
| Vertebrates Home | Vertebrate | Vertebrate |
|
Abbreviated Dendrogram
Diapsida
├─Archosauromorpha
│
└─Lepidosauromorpha
├─Sauropterygia
│ ├─Placodontia
│ ├─Pachypleurosauridae
│ └─┬─Nothosauridae
│ └─Plesiosauria
│ ├─Pliosauroidea
│ └─Plesiosauroidea
│ ├─Elasmosauridae
│ └─Cryptocleidoidea
└─Lepidosauriformes
├─Sphenodontia
└─Squamata
|
Contents
Overview |
Life reconstruction of the Early Jurassic necked plesiosaur Plesiosaurus dolichodeirus in underwater flight. Drawing by Dr Adam Stuart Smith (Plesiosauria com), copied from Wikipedia, Creative Commons Attribution Share-Alike |
There is some controversy as to how these creatures moved, as no living sea animal propels itself with both pairs of flippers [with the possible exception of Latimeria, which has five flippers, including a highly modified anal fin]. The original notion was that plesiosaurs "rowed" like an ancient galley. Based on functional studies of plesiosaur limbs, Robinson (1975, 1977) argued that plesiosaurs swam using their paddles in an up-and-down in a "flying" motion to propel themselves through the water, as Recent marine turtles do. The paddles thus acted like hydrofoils generating thrust during the backstroke and also from lift during the recovery stroke. This hypothesis has since been modified, and the current view is that plesiosaur's swam much like modern sea lions. To appreciate the problem, we need a bit more understanding of the plesiosaur limb girdles.
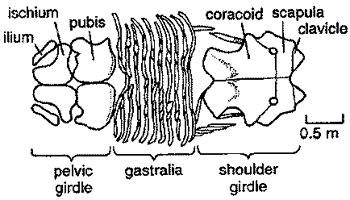 The basic scheme is shown in the figure at right, modified from Taylor 1981) [1]. The pelvic girdle is similar to the basal sauropterygian pattern. However, the shoulder girdle has been highly modified from the ancestral pattern discussed previously. Anteriorly, the clavicles are no longer the main support, and the plate-like scapulae are in broad contact along the midline. Carroll & Gaskill (1985) have examined growth series for Plesiosaurus and have shown that the juvenile arrangement is almost indistinguishable from the basal sauropterygian pattern. With increasing size, the scapulae grow towards each other. Eventually, they contact each, filling the space between the two sides.
The basic scheme is shown in the figure at right, modified from Taylor 1981) [1]. The pelvic girdle is similar to the basal sauropterygian pattern. However, the shoulder girdle has been highly modified from the ancestral pattern discussed previously. Anteriorly, the clavicles are no longer the main support, and the plate-like scapulae are in broad contact along the midline. Carroll & Gaskill (1985) have examined growth series for Plesiosaurus and have shown that the juvenile arrangement is almost indistinguishable from the basal sauropterygian pattern. With increasing size, the scapulae grow towards each other. Eventually, they contact each, filling the space between the two sides.
Posteriorly, the coracoids are more massive and are elongated posteriorly to form a continuous ventral shield. During development, the coracoids both send strong midline processes forward until they meet the scapulae, splitting the vacuity at the center of the girdle. Ultimately, all that is left of the formerly large hole at the center of the pectoral girdle are two small coracoid foramina, as shown in the figure at right.
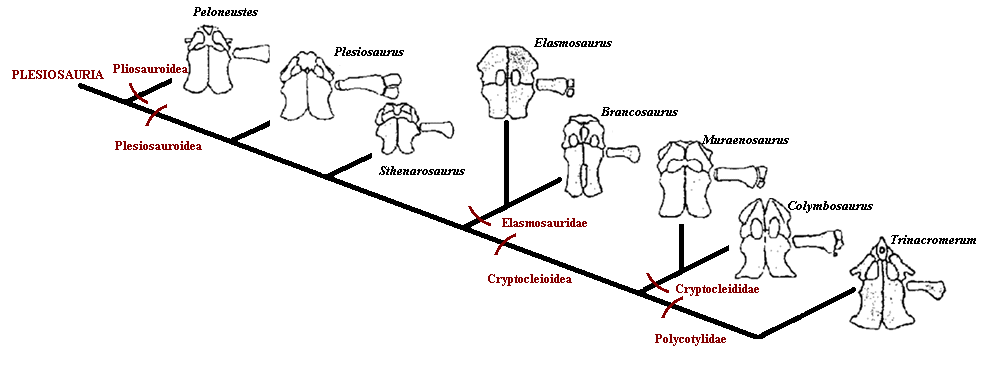
Because the shoulder and ventral girdles are so significant a part of plesiosaur anatomy, they are often very distinctive in different species, and are a useful way of identification and classification of this group. The evolution of shoulder girdle is shown against the overall scheme of plesiosaurian evolution in the figure below.
The gastralia, found in some pachypleurosaurs, are expanded in plesiosaurs to become a massive mesh of ventral ribs. These were presumably tightly linked to each other and to both sets of limb girdles by stout ligaments. Since all of the main limb girdle elements were in broad medial contact, they were no longer free to flex laterally or to rotate with respect to each other. Accordingly, a strong but flexible ventral "spring" was required to allow the animal to move at all. In essence, the shoulder and hip girdles were suspended within the ventral basket which was held under the vertebral column like the string in an archer's bow. This elastic suspension could absorb the up-and-down motion of the limb girdles at each beat of the limbs, while transmitting the propulsive force to the body. Robinson (1975, 1977).
Thus, we may model the shoulder and hip girdles of plesiosaurs as if they were broad, flat bony plates with limb joints on opposite edges. The main propulsive muscle masses lay on top and under the girdles, and their tendons attached to the dorsal and ventral surfaces of the limbs and swept them powerfully in the desired directions. Supplementary muscles controlled the trim of the flippers so that on both up- and down-stroke the hydrofoil surfaces of the limbs, moving in the vertical plane, generated a resultant thrust forwards. Effectively, therefore, each pair of limbs mounted on its girdle was a self-contained power unit producing a forwardly directed propulsive force.
These two power modules were supported within the domed body, by a distinctive 'ventral basket' made up from the two limb girdles bound together by ligaments running onto and between the gastralia, the splint-like bones of the middle abdominal floor. Robinson argues that this acted as a string to the 'archer's bow' formed by the vertebral column. The contraction of the muscles between the dorsal processes of the vertebrae bent the ends of the column upwards and pulled taut the ventral basket suspended from the vertebral column by its ligamentous attachments. With the ribs, this formed a remarkable suspension system restraining the girdles while allowing some elasticity to absorb the up-and-down motion of the girdles with each beat of the limbs. The ability to vary independently the action of each limb allowed rapid change of direction in the pursuit of prey. The arched vertebral column might even allow plesiosaurs to crawl up on land to breed without collapsing the lungs, like modern sea turtles and unlike Recent whales or the contemporary ichthyosaurs.
Although the functional anatomy of the plesiosaurs makes good sense overall, it is not as clear exactly how the plesiosaurs exploited the locomotor possibilities of their unique body plan. Their distinctively domed body, with its four large broad-bladed limbs, has no precise modern ecological counterpart. It is the old problem of trying to figure out the lifestyle of a creature that has been dead for tens of millions of years.
Unlike other marine reptiles, Plesiosaurs have relatively small tails, but large and powerful paddles, so it is assumed that the latter were used in creating thrust. Exactly how they did is the problem. Three possible hypotheses have been proposed. These are:
 |
 |
 |
| Rowing underwater like a duck, with an active power stroke backwards (bold line) then a recovery stroke forwards (dotted line). The plesiosaur's limb shape is particularly unsuited to such a movement. | Flying underwater like a turtle or penguin, with an up-and-down beat of the limbs; this movement would have been ideal for the limb shape but only the downstroke could have been really powerful. | Swimming like a sea lion, with a backwards power stroke and forwards recovery stroke, a movement combining elements of the other two modes |
The lines indicate the movement of the tip of the flipper.
For a long time scientists thought that plesiosaurs moved along by simply rowing their long limbs back and forth. This would seem the most obvious explanation. In rowing, the limb sweeps backwards, while its vertically aligned blade bites into the water and levers the animal forwards. The limb then returns forward to start another power stroke. A limb which is adapted to rowing therefore has to be rigid and oar shaped, broadening at the end, and moves in horizontal plane.
With plesiosaur paddles, this mode of locomotion would not work because the recovery stroke would create a counter-thrust which would oppose the forward motion to some extent, since the paddles could be feathered (i.e. tipped to the horizontal), but not removed from the water. If plesiosaurs really had rowed through the water, they should have had webbed folding feet like those of ducks. This is obviously not the case. As pointed out by Robinson, the limb of a plesiosaur is of a quite different design and has characteristic adaptations to undersea flight. Compare the limbs of Ceresiosaurus, which seem better designed for this duck-like locomotion.
Robinson (1975) argued that the plesiosaurs must have "flown" underwater, like sea turtles and penguins, because this movement was the most efficient use of their long hydrofoil-like limbs. She suggested that they swam by flapping both pairs of limbs up and down, producing lift force that propelled the animal forwards. The hydrofoil surfaces swept up and down producing lift forces which averaged out over the whole cycle to a forwardly directed propulsive resultant force
When we look at the plesiosaur limb, we see that it is flat with an aerofoil cross-section like a bird's wing. Or, more correctly, a hydrofoil, because we are speaking here about a "wing" that "flies" through the water.
The plesiosaur limb has a specific shape - it tapers towards its tip to reduce drag, unlike the oar-like shape of the rowing limb. The plesiosaur limb also has just enough flexibility to allow trimming and to damp out drag generating eddies. This arrangement is, again, quite unlike the rigid rowing limb which relies on its production of drag to bite into the water and gain leverage. In fact the limbs of plesiosaurs are similar to the forelimbs of marine turtles, sea lions and penguins, all of which all act as hydrofoils.
The cross-section is such that when it moves through the water with the plane of the limb edge-on to the direction of movement through the water it generates lift (it is the same principle as a bird or airplane wing generating lift in the air). This lift is perpendicular to the direction of movement of the limb. If such a paddle pushes down and backwards at an angle above the horizontal, it generates lift and forwards thrust. In the upstroke, the paddle is tipped the other way, and a further smaller forwards lift force is generated. The paddle tip describes a figure-of-eight pattern and each stage of the cycle produces forwards movement. The plesiosaur limb, in this hypothesis, has to move up and down in the vertical plane to produce horizontally forward directed propulsive force
In 1982 Tarsitano & Riess suggested a modification to Robinson's underwater flight hypothesis. Although agreeing that plesiosaurs must have used their limbs to fly underwater, they noted that the simplest and apparently most effective up-and-down action was anatomically impossible. The bones of the shoulder joints would block movement of the forelimbs above the horizontal plane. Moreover, plesiosaurs do not seem to have had an equivalent to the avian triosseal canal which would allow muscles located in the trunk to pull the flipper above the muscle's own horizontal level.
In the same year Frey & Riess proposed that the 'wings' had an active downstroke, and a comparatively passive upstroke as they returned to the starting point. They suggested that the two pairs of limbs beat out of phase: one pair propelling the animal during its down stroke while the other was brought back to the start of its own power stroke. This limb action would be relatively inefficient compared with that of modern animals such as penguins, whose wings produce propulsive force throughout virtually the whole of the upstroke and downstroke.
Two years later, Godfrey 1984) suggested a different modern analogy for plesiosaur locomotion: sea lions. The sea lion swims by beating its forelimbs forcefully downwards and then relatively passively upwards, while always moving them backwards. Once fully retracted, the limbs can then be feathered and brought forwards during a recovery stroke. In this modified version of the flying model, the paddle tip describes a crescent-shaped path as in the sea lion.
His argument is based on the anatomy of the plesiosaur skeleton. Plesiosaurs could not have moved their paddles up and down in a figure-of-eight because the pectoral and pelvic girdles are both flattened heavy units of bone that form an immovable ventral bony plate. The gastralia between the girdles are too weak for strong vertical movements. Evidence does however point to the plesiosaurian shoulder and hip girdles having the massive muscles needed for the limb action characteristic of sea lions.
Interestingly, sea lion swimming combines elements of both underwater flight up-and-down action) with rowing (backwards and forwards movement). This appears to be the most plausible explanation for plesiosaur swimming. MAK990911, ATW020630
[1] In the original figure, the labels for scapula and coracoid appear to be reversed. This has been "corrected" here, but the reader is warned to be cautious.
checked ATW050109, revised MAK111126
Using this material. All material by ATW is public domain and may be freely used in any way (also any material jointly written by ATW and MAK). All material by MAK is licensed Creative Commons Attribution License Version 3.0, and may be freely used provided acknowedgement is given. All Wikipedia material is either Gnu Open Source or Creative Commons (see original Wikipedia page for details). Other graphics are copyright their respective owners
{kind=link}