 Pythonomorpha:mosasaurs + snakes
Pythonomorpha:mosasaurs + snakes| Squamata | ||
| The Vertebrates | Pythonomorpha |
| Vertebrates Home | Vertebrate | Vertebrate |
|
Abbreviated Dendrogram
Squamata
├─Iguania
└─Scleroglossa
│
└─Pythonomorpha
├─┬─Aigialosauridae
│ └─Mosasauroidea
└─Serpentes
├─Scolecophidia
└─Alethinophidia
├─Aniloidea
└─Macrostomata
├─Boidae
└─Caenophidia
├─Acrochordoidea
└─Colubroidea
├─Colubridae
└─Viperidae
|
Contents
Overview |
Lee (1997a) is one of those rare papers that almost immediately convinced most paleontologists of its central proposition: that snakes are more closely related to mosasaurs than to varanid lizards. Varanids are represented today by monitor lizards, gila monsters, and a few close relatives. The extant varanids are all big, active hunters (at least by lizard standards). By contrast, mosasaurs were almost all gigantic aquatic predators who flourished in the Late Cretaceous and apparently died out with the dinosaurs. For many years, the orthodoxy was that snakes were highly derived varanids of some sort. However, as early as 1869, Edward Cope had argued that snakes were more closely related to mosasaurs, and he coined the term "Pythonomorpha" to describe the group that united them. This idea was occasionally also floated by others, but remained essentially dormant for almost five generations.
Michael Lee learned cladistic analysis in a rugged school. His doctoral work in England in the early 1990's concerned the origin of turtles. He advanced the idea that turtles are living pareiasurs. This hypothesis was, and remains, highly controversial; and Lee was forced to fight epic battles with unbelievers, culminating in two extensive, back-to-back articles by Lee (1997) and by deBraga & Rieppel (1997) in a single issue of the Zoological Journal of the Linnean Society.
By the time this work was published, Lee was already back in Australia preparing his next major project: the revival of Cope's pythonomorph concept. He attacked this project with characteristic energy and with his trademark attention to obscure fossil specimens. The work was published in the same year as the ZJLS article, but received a very different reception. As one paleontologist noted only a year later, "most of the workers I've spoken to that are aware of this work support it virtually 100%." It is truly a beautiful piece of work. Unlike most cladistic works, Lee provides figures for most of the characters he describes, which is enormously helpful for the 99% of his readers who are not lizard specialists. He carefully analyzes fabulously obscure specimens like Saniwides and Paravaranus. He explicitly states his higher-level phylogenetic assumptions. Almost the only element short of perfection is that, hidden in a mass of detail, are a substantial number of uncertain judgment calls about the scoring and polarity of certain characters. However, the end result is a convincing case that Pythonomorpha is a very strongly supported clade.
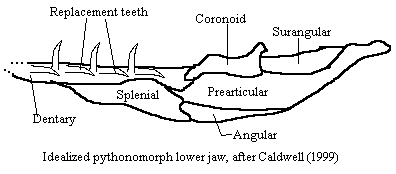
In fact, oddly enough, Pythonomorpha and its component clades were some of only a handful of clades which were strongly supported. Most of the relationships among the Varanoidea disappeared into vague polyotomies when bootstrapping tests were run. That is, their existence and position was very sensitive to minor variations in the data. Thus, Lee's use of poorly known taxa and the large number of assumptions made about higher level phylogeny and polarity make the rest of his cladogram fairly suspect. However, Pythonomorpha was extremely robust, and Lee identified 38 synapomorphies of the group, many of which dealt with the peculiar mobile lower jaw figured in the main entry in these Notes.
This conclusion was verified by Caldwell (1999), who used a rather different approach. To say "verified" is somewhat unfair, since Caldwell was working independently on the question and submitted his original manuscript three months before Lee did. However, the paper was held up in review for a year and in press for two more. Caldwell's primary work has always been in this phylogenetic area, particularly with aigialosaurs. Aigialosaurs are generally thought to be the sister group of mosasaurs, or may even include mosasaurs. Caldwell was once of the view that aigialosaurs were basal, largely terrestrial varanoids, probably close to snakes, and that mosasaurs were, at least morphometrically, something quite different. Caldwell et al. (1995). However, he seems to have come around to Lee's take on Pythonomorpha about the same time as Lee did.
As with Lee's work, Caldwell's 1999 study strongly supports Pythonomorpha. This is particularly informative because Caldwell's study is considerably different. He makes few assumptions about higher-level lizard taxonomy and includes gekkos, skinks and agamids among his ingroup taxa. He uses fewer of the poorly known fossil species and concentrates on well-characterized living groups, which minimizes the amount of missing or ambiguous data (at the risk, of course, of missing key transitional forms). His outgroup is Sphenodon, so there are no questionable assumptions about polarity. Caldwell also holds some rather different views about homology. He is far less likely than Lee to take osteological data at face value, and he attempts to exclude characters a priori when he perceives them as supporting fundamentally different structures.
Interestingly, Caldwell also used the ACCTRAN option in PAUP. ACCTRAN and DELTRAN are two different optimizing strategies. ACCTRAN assumes, whenever the results are ambiguous, that character transitions happen at the earliest possible point. Thus ACCTRAN tends to generate synapomorphies of higher level taxa and favors reversals over convergences. DELTRAN takes the opposite approach. DELTRAN therefore tends to generate apomorphies of terminal taxa and favors convergences over reversals. [1] He also seems not to have had access to a particularly good computer, as he was apparently forced to use the "heuristic" or step-wise option of PAUP. With this option, taxa are added one at a time, in random order, and tacked on to the tree as they are added. This can result in slightly different trees depending on the order of addition. The heuristic option therefore does not ensure a global most parsimonious result and requires many repetitions to avoid missing the optimum path. Possibly as a result of these constraints, Caldwell's strict consensus tree is one enormous polyotomy -- considerably less dramatic than Lee's result. However, application of the "majority rule" easily produced the same result as Lee's paper as far as the Pythonomorpha were concerned.
Not surprisingly, Caldwell's results outside Pythonomorpha are markedly different from Lee's. Lee constrains the Pythonomorpha to fall among the varanoids. Caldwell concludes that varanoids and pythonomorphs could not possibly be further apart without living on different planets. However, both of these results are scientifically insignificant. If anything, Caldwell's results demonstrate that the basal lizard polyotomy is even less resolvable than previously thought. However -- and this is the important thing -- Caldwell and Lee both find strong support for Pythonomorpha: the close relationship between mosasaurs and snakes.
Thus, Pythonomorpha is supported by two, quite divergent studies, which is as close to proof as one is likely to find in this area. Caldwell's higher-level taxonomy has been incorporated in these Notes, but the choice is relatively arbitrary. It is very uncertain where the ancestral pythonomorph derived from. However, somewhere in the early Cretaceous, a medium-large, low-slung, long-bodied, probably terrestrial predacious lizard developed a unique hinge in the middle of its lower jaw, loss of the symphysis joining the two halves of the lower jaw, and a unique way of stabilizing the spinal column (the zygosphene-zygantra articulation). These features (all of which were among those noted by Cope in 1869!) seem to have been a major evolutionary success in a number of quite different environments and may have driven the successive radiations of aigialosaurs, mosasaurs, and snakes, the last of which continues today. --ATW 000723
[1] I understand, thanks to a careful reader, that ACCTRAN is by far the more usual option. This would appear to be the more conservative approach. However, the fact remains that it promotes the formation of terminal polyotomies because the higher level nodes "soak up" the available synapomorphies, particularly if there is significant missing data. Suppose, for example, that taxa a, b & c are scored for "purple hair". Taxon 'c' has purple hair but 'a' and 'b' are fossils with no known hair. Under ACCTRAN, the node ABC will be scored as being supported by the synapomorphy "purple hair." Under DELTRAN, this character will be an apomorphy of taxon 'c'. If 'a' and 'b' are otherwise fairly similar, we may see a polyotomy at ABC under ACCTRAN, as well as a list of synapomorphies for ABC which is inflated by the addition of a characteristic which is in fact unknown. Normally, this is harmless. Sometimes, it is not. Back --ATW 011226.
According, to the Biblical literalists, the Serpent was responsible for getting Adam and Eve to try Apples. As punishment, the snake lost his legs, and the human race was burdened with the Windows OS. For all its obvious failings, this tale provides almost as much scientific satisfaction as the current state of research in the area.
This essay is, chronologically, the fourth attempt in these Notes to make some sense of early snake evolution. See the essays related to the Scolecophidia, Pythonomorpha, and Aniliidae. Many of the positions taken in these essays, all written within the last two years, are already pretty clearly wrong. The Aniloidea, for example, appear to be monophyletic; and the scolecophidians probably are a sideshow to the main line of snake evolution, after all. Having been steered wrong so often before, you may well ask why you should be getting on this particular bus again. The answer is that today we will not be attempting to actually go anywhere. Instead, this will be a simple sightseeing trip -- albeit one into the middle of a war zone.
The contending forces are familiar names. Arrayed on one side are Michael MSY) Lee and Michael Caldwell, with a cast of numerous others. They contend for a marine, perhaps mosasauroid origin for snakes. Against them stand the Rieppel People, mostly Olivier Rieppel from the Field Museum and Hussam Zaher from the University of São Paulo (via the AMNH). They contend that Lee's marine snakes were already derived Macrostomates and that snake origins lie buried in the earth, among fossorial lizard-kind.
For a moderately obscure issue, this controversy has generated a remarkable amount of heat. Perhaps the story goes back to the disagreement between Rieppel and Lee concerning the relationship of turtles and pareiasaurs. The first sign of trouble on the snake front actually began before the pareiasaur matter wound down. In 1996, both Lee and Caldwell submitted papers supporting mosasaurs as at least) the sister group of snakes. Lee (1997a); Caldwell (1999). While these papers were in review, the two collaborated on redescribing a possible intermediate form, an enigmatic "varanoid lizard" which had been gathering dust at the Hebrew University in Jerusalem since the mid-1970's. This specimen (actually two specimens) was Pachyrachis, which turned out to be a very early marine snake. Caldwell & Lee 1997). Given the mosasauroid connection and the marine nature of Pachyrachis, Caldwell & Lee proposed that the snakes evolved from marine forms, rather than fossorial lizards. The two presented a very thorough cladistic analysis which appeared to support this hypothesis.
Just two months later, Hussam Zaher wrote and submitted a short rebuttal piece, which was published as a "Rapid Communication" in JVP the following year. Zaher (1998). Zaher argued that Pachyrachis was a derived, macrostomate snake so that its marine character proved nothing about the origin of snakes. Zaher, Rieppel and others then discovered another forgotten Israeli marine specimen from Hebrew University, Haasiophis, which they incorporated in a cladistic analysis which supported their view of the matter. Tchernov et al. (2000).
Caldwell replied to Zaher's piece in JVP in 2000. Caldwell (2000). Meanwhile, Lee literally went to the opposite ends of the earth. In Australia, he redescribed Wonambi, perhaps the only decently preserved madtsoiid snake in existence, and incorporated it into the Caldwell-Lee cladogram. Scanlon & Lee (2000). To the surprise of absolutely no one, the results confirmed their earlier conclusion that snakes came of marine ancestry. To complete the circle, I made a rough pass at coding Wonambi into the Zaher-Rieppel character matrix and ran it on PHYLLIP (the poor man's PAUP). It will shock no reader to hear that the results supported the Zaher-Rieppel terrestrial origin hypothesis.
So what is one to make of all this? Truthfully, its hard to say. These Notes reflect the Zaher-Rieppel model for now, but that choice was fairly arbitrary. The Caldwell-Lee character matrix is vastly more complete, and the analysis of post-cranial characters is incomparably better. But it is structured in a way that raises a number of questions to which there are no immediate answers.
For one, the number of dependent characters is striking. In the Wonambi paper, by way of example, character 97 is "Ectopterygoid. 0: large. 1: small. 2: absent." Characters 98, 99, and 100 then relate to various contacts and conformations of the ectopterygoid. Obviously if character 97 is coded as state '2' (ectopterygoid absent), characters 98-100 are rather meaningless. The note for character 99 states that it cannot be coded for animals without an ectopterygoid. Does this mean it is treated as missing data, as an additional character state of "unknown," or in some other way? What about characters 98 and 100? To make matters worse, character 100 is also expressly dependent on an uncoded state of the jugal, and the jugal itself is absent in advanced snakes (see character 44). Confused? It gets worse.
Most likely, the "not applicable" codings are treated as missing data, as this is the preferred method. What happens to missing data? Under PAUP, missing data can take any value and is treated so as to assume whatever value gives the smallest number of character changes. So the question is like asking, "what color is a chameleon?" It takes on the "color," or in this case the ectopterygoid, of its background. For anything but really small data sets, PAUP adds data stepwise, one taxon at a time. It draws the best tree it can from those data, then adds the next taxon. The order of addition is typically randomized and the process is repeated a few hundred times to find the shortest trees. (This is an oversimplification, but not too far off.) Now, consider what happens if we have a small number of taxa with lots of missing data: Pachyrachis, Wonambi, and Dinilysia for example. Most of the other taxa are living snakes with complete data sets. The first fossil taxon to be added will take on the ectopterygoid of the living example closest to it in other respects. Well and good. That's the way its supposed to work. But the second fossil taxon -- providing it is not too radically different from the first -- will be "attracted" twice as strongly to first because, in this way, it can minimize its distance from two other taxa. Thus, generally similar fossil taxa will tend to clump if they are mixed in with lots of living taxa. Also, taxa, living or fossil, with absent bones (e.g., without an ectopterygoid) will tend to clump in a data set having lots of snakes with an ectopterygoid if there are many dependent characters.
To make matters worse, Scanlon & Lee (2000) use soft tissue data -- lots of it. Characters 192 to 234 are all soft tissue, and thus all missing data for the three fossils. The soft tissue data set that they use is mostly that of Cundall et al (1993). This is discussed in connection with the anilliids. You may recall that Cundall's group was quite cautious about this data because, by itself, it resulted in phylogenetic gibberish. Osteological data alone showed a monophyletic Aniloidea. When combined with the soft tissue data, the total data yielded a paraphyletic Aniloidea, a result we now believe to be wrong. This suggests that convergent evolution is at work in the soft tissue characters. The legless lifestyle, after all, poses similar problems for all snakes, and there are only so many solutions. Since muscles seem to change much faster than bones, Cundall's results could be explained by invoking convergence and perhaps long branch attraction. This would tend to break up the monophyly of the aniloids by dragging some members closer to the macrostomate species which they most closely resemble by convergence and chance. The addition of fossil taxa compounds the problem of phenetic convergence because the misleading convergent soft tissues could create a completely erroneous phylogenetic center of gravity at which the fossil taxa would clump due to the large amount of missing data.
There's lots more, but the reader's patience for cladobabble is probably limited. None of these comments are intended as necessarily precluding the marine origin of snakes. Far from it! However, the Caldwell-Lee data set raises a number of issues that need to be addressed. Fortunately, these are the kinds of questions which can be answered by anyone with a reasonably good computer and a few days to spare. Perhaps we will do so ourselves the next time we return to this pit of contending vipers. ATW 010902
References: Caldwell 1999); Caldwell (2000); Caldwell & Lee (1997) [C&L]; Cundall et al. (1993); Hecht (1959); Lee 1997a); Lee et al. (1999) [LCS]; Lee et al. (1999a) [LBC]; Scanlon & Lee (2000); Tchernov et al. (2000); Zaher (1998). 010826.
Pythonomorpha:mosasaurs + snakes
from the Late Cretaceous.
Scleroglossa: (Amphisbaena + Gekkota) + (Scincomorpha + Anguimorpha)) + *: (Mosasauroidea + Aigialosauridae) + Serpentes.
$ Recumbent replacement teeth; thecodonty; $ four or fewer teeth on premaxilla; $ mandibular symphysis absent (i.e. sides of the lower jaw are not firmly joined anteriorly); $ vertical, mobile articulation of splenial and angular, allowing bulging of lower jaw (reversed in most scolecophidians) [LBC]; dentary overlap with post-dentary bones reduced or absent; splenial may be reduced or with reduced sutural contact with dentary; splenial does not overlap? contact? coronoid; coronoid articulates with dentary; supratemporal contacts prootic; posterior ramus of coronoid absent; septomaxilla with long posterior process (may reinforce central axis of elongated upper jaw); pterygoid teeth present; palatine with long anterior process; reduced basipterygoid process; reduced interpterygoid vacuities; parietal with large descending flange sutured to prootic; supratemporal contacts prootic; ribs begin with third cervical; $ zygosphene-zygantra articulations in vertebral column; pelvic elements not sutured; femur stout (if present); osteoderms absent.
Links: Mosasaurs; Mosasauridae Translation and Pronunciation Guide (see entry for "Pythonomorpha"); Squamata. Note: For further information on this taxon, the Oceans of Kansas web site has a great deal of specialized information on mosasaurs, perhaps the most typical of pythonomorphs; Pythonomorpha (Mikko's Phylogeny); Molecular evidence for a terrestrial origin of snakes reprint of Vidal & Hedges (2004) with a molecular study, also finding snakes outside of Varanoidea); The braincases of mosasaurs and Varanus, and the relationships of ... same finding).
Note: Lee lists 38 synapomorphies of this group. Only a few are detailed here.
References: Caldwell 1999); Caldwell et al. 1995), deBraga & Rieppel 1997), Lee (1997), Lee (1997a); Lee et al. (1999a) [LBC] ATW040704.
 Aigialosauridae:Aigialosaurus, Carsosaurus, Coniasaurus, Opetiosaurus.
Aigialosauridae:Aigialosaurus, Carsosaurus, Coniasaurus, Opetiosaurus.
Late Jurassic (Tithonian) to Late Cretaceous (Turonian) of Europe.
Pythonomorpha:: Mosasauroidea + *.
Notes:paraphyletic?
Links: The Journal of Vertebrate Paleontology; Untitled Document; The Journal of Vertebrate Paleontology; 2002 The 15th Tokyo .... ATW030330
upK.
Pythonomorpha:: Aigialosauridae + *.
mesokinetic fronto-parietal hinge & metakinetic parietal-occipital hinge partially eliminated by tight overlaps but not closed sutures [LBC]; anguiliform motion w. limbs for steering; 10m; shallow marine; differ from Varanoidea in fusion of frontals, extension of premaxilla to frontals; joint in lower jaw; teeth specialized for various marine prey; post-cranial skeleton modified for aquatic motion & large size.
Links: Oceans of Kansas Paleontology (Obviously Best on the Web); The DOL Dinosaur Omnipedia; Mosasauroidea after Bell, 1994.
References: Lee et al. (1999a) [LBC].
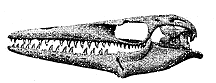
Image: Clidastes velox, originally from Williston, SW (1898), Mosasaurs, in The University Geological Survey of Kansas, Volume IV, Paleontology. State Printer of Kansas, V(? or I?): 81-347, and reprinted at Mike Everhart's incomparable Oceans of Kansas site. Note: I have made no serious attempt to characterize this taxon, and gladly defer to the Oceans of Kansas. I would add only that mosasaurs are large, easily studied organisms known from many, beautifully preserved and relatively complete specimens almost worldwide. Anyone interested in vertebrate structure and function should pay particular attention to mosasaurs for that reason, as well as their marine adaptations and key phylogenetic position. Mosasaurs are relatively basal squamates but, at the same time, close relatives of snakes, the most derived squamates.
Serpentes: ~Ophidia): snakes. Since these Notes currently have nothing snaky which is more basal than the crown group of snakes, Ophidia and Serpentes are treated as synonymous. Actually, Ophidia = mambas > mosasaurs; and Serpentes = boas + blindsnakes. In these notes, Serpentes is used as a crown group, with Scolecophidia and Alethinophidia treated as complementary stem groups.
fr upK.
skull highly kinetic, with up to 8 links permitting rotation or expansion, including lower jaw joint; advanced forms have essentially no rigid skull except braincase; $ marginal teeth ankylosed to to rims of discrete sockets [C&L]; teeth frequently angulated rather than smoothly recurved [LCS]; $ dentary with <3 mental foramina [C&L]; compound bone present [C&L]; at least some mobility of maxilla and palatal arch [LBC]; $ palatines with distinct medial rectangular process [C&L]; chemosensory vomeronasal "Jacobson's" organ in roof of mouth -- tongue used to sample air, transport to organ; $ premaxilla-maxilla articulation nonsutural and mobile [C&L]; lacrimal absent [C&L]; eyes permanently covered by transparent lower lid in most modern snakes; eyes: lens is moved, not bent, to focus; no fovea; no oil droplets; no sclerotic support; unique double cone (suggests ancestor was possibly blind cave, marine or burrowing form); mesokinetic fronto-parietal hinge eliminated [LBC]; frontal and parietals completely enclose brain; metakinetic parietal-occipital hinge eliminated [LBC]; supratemporal superficial to parietal [C&L]; jaw adductors "insert" (originate?) on dorsal surface of parietal [C&L]; parietal table reduced to sagittal crest [C&L]; $ descending process of parietal contacts parabasisphenoid [C&L]; no tympanum or middle ear; $ tympanic recess absent [C&L]; some detection of vibration through lower jaw and skin at low frequency (<1K cps); $ >140 precaudal vertebrae [C&L]; vertebrae proceolous, with well-developed zygosphenes, zygantra, diapophyses; hypapophyses usually present in some region of spine; $ pectoral girdle & limbs absent [C&L]; pelvic girdles, limbs rudimentary or absent; numerous specializations related to elongation & locomotion, e.g., loss of 1 lung (but elaborations of trachea may functionally serve as additional lung), 1 of paired gonads & kidneys displaced longitudinally.
Links: ANATOMY OF A SNAKE; The EMBL Reptile Database; Squamata/ophidia01.html">Reptilia, Squamata, Ophidia - Schlangen (German); Ophidia -Faszination Schlangen- (German); The Ophidia Files; A revision of the Australasian Pythons - Hoser - 2000..
References: Caldwell 1999); Caldwell (2000); Caldwell & Lee (1997) [C&L]; Cundall et al. (1993); Hecht (1959); Lee 1997a); Lee et al. (1999) [LCS]; Lee et al. (1999a) [LBC]; Scanlon & Lee (2000); Tchernov et al. (2000); Zaher (1998). 010826.
checked ATW060222