 The Classical View
The Classical View| Archosauria | ||
| The Vertebrates | Archosauria |
| Vertebrates Home | Vertebrate | Vertebrate |
|
Abbreviated Dendrogram
Archosauromorpha
│
└─Archosauria
├─Proterosuchidae
└─┬─Erythrosuchidae
└─┬─Euparkeriidae
└─Crown Group Archosauria
├─Ornithodira
│ ├─Pterosauria
│ └─Dinosauromorpha
└─Crurotarsi
├─Phytosauridae
└─Rauisuchiformes
├─Rauisuchia
│ ├─Ornithosuchidae
│ └─┬─Prestosuchidae
│ └─Rauisuchidae
└─Suchia
├─Aetosauridae
└─Crocodylomorpha
|
Contents
Overview |
Archosaurian systematics have sometimes been a contentious area. It would be unproductive to relate the entire history of the field. However, it is important to have a rough understanding of the systematic frame of reference used by many of the older papers.
The Classical ViewBy the middle years of the last century, The term "archosaur" had come to include crocodiles, dinosaurs, pterosaurs and a wide variety of "primitive" Triassic reptiles. Generally speaking, it would include what we would now call Archosauriformes, except for birds. The more basal archosauromorphs were not recognized as part of this lineage. At that time, rhynchosaurs, for example, were thought to be part of the "Lepidosauria," which encompassed some basal Diapsida ("Eosuchia") as well as Lepidosauriformes. Other archosauromorphs were classified with the "Euryapsida," a vague group which swept in most of the Mesozoic marine reptiles as well as others, such as Trilophosaurus, which were thought to be related. The recognized higher orders of Archosauria were Crocodilia, Pterosauria, Saurischia and Ornithischia (the monophyly of dinosaurs was not well established). This left a large number of Triassic species in a paraphyletic group of basal archosaurs, the "Thecodontia." The thecodonts, in turn, were divided into three suborders -- essentially perceived as evolutionary grades on the way to the crocodiles: Proterosuchia (basal Archosauriformes), Pseudosuchia (Crurotarsi except Phytosauridae and Crocodylomorpha), and Parasuchia (Phytosauridae). While it was believed that dinosaurs and pterosaurs derived from the thecodonts, no one had a good idea of where the divergence had occurred. Perhaps the most popular theory was that the dinosaurs and pterosaurs were the result of multiple radiations from several different thecodont lines. See, for example, Walker (1964).
 During the 1970's and 1980's, two developments took place that made a fair mess of the old system and fractured the paleontological community: the advent of cladistic methods and the early anatomical and locomotor studies on the archosaur tarsus (ankle) by Krebs, Cruikshank, Chatterjee, and others. Now, a generation later, younger scientists are writing review articles which seem to wonder what the fuss was all about. See, for example, Brochu (2001) and Gower & Wilkinson (1996). From today's point of view, it appears that everyone was reaching about the same conclusions at about the same time. For example, by the mid- or late 1980's almost everyone agreed that: (a) there was a single main split between the croc lineage and the dino-pterosaur line; (b) Euparkeria was very close to that split; (c) forms we now consider to be basal archosauromorphs were just that; (d) Scleromochlus was on the dinosaur branch; (e) almost all of the other known "thecodonts" beyond Euparkeria were on the croc side of the split; and (e) ornithosuchids and phytosaurs were fairly basal archosaurs. That's obviously very broad agreement. Why all the hard feelings? Perhaps it isn't science, but the question deserves an answer.
During the 1970's and 1980's, two developments took place that made a fair mess of the old system and fractured the paleontological community: the advent of cladistic methods and the early anatomical and locomotor studies on the archosaur tarsus (ankle) by Krebs, Cruikshank, Chatterjee, and others. Now, a generation later, younger scientists are writing review articles which seem to wonder what the fuss was all about. See, for example, Brochu (2001) and Gower & Wilkinson (1996). From today's point of view, it appears that everyone was reaching about the same conclusions at about the same time. For example, by the mid- or late 1980's almost everyone agreed that: (a) there was a single main split between the croc lineage and the dino-pterosaur line; (b) Euparkeria was very close to that split; (c) forms we now consider to be basal archosauromorphs were just that; (d) Scleromochlus was on the dinosaur branch; (e) almost all of the other known "thecodonts" beyond Euparkeria were on the croc side of the split; and (e) ornithosuchids and phytosaurs were fairly basal archosaurs. That's obviously very broad agreement. Why all the hard feelings? Perhaps it isn't science, but the question deserves an answer.
The hard feelings were generated precisely by differences in methodology. The traditional taxonomist says "I have studied these bones for 30 years. I have the judgment, experience and wisdom to see patterns and distinctions which you cannot, and so you should listen to me." The cladist says: "I have concentrated on point by point comparisons of dozens of specimens. My conclusions are the result of truly objective computer-based analysis -- not mere intuitions, and so my view is the only scientific one." The anatomist says: "All of you ignore these animals as living organisms. I have studied the detailed relations between the bones. Only I consider how the animal worked as a living organism, and this is the only way one can understand the evolutionary mechanisms at work."
Of course, all three of these hypothetical folks are correct -- except for their claims to exclusivity. Thus, it is not surprising that all three groups were reaching the same conclusions. However, notice that each group's attitude could be perceived as not just disagreeing with some trifling discrepancy in the others' results, and not just disagreeing on methodology, but as attacking the others personally. The taxonomist is heard to question the judgment and maturity of the cladist and anatomist. The cladist may be implying that the others are not objective -- and not even scientific. The anatomist may be implying that the cladist and traditionalist don't really know anything useful. Scientists being no better and no worse than other human beings, sometimes they did mean to engage in personal attacks. Mostly they did not. However, the three methodologies of that generation represented very different world views, and the inevitable philosophical conflicts sometimes overshadowed scientific agreement.
 The Code War: the Clash of the Cladograms
The Code War: the Clash of the CladogramsAs the Cold War was ending, the Code War was just heating up. The first serious attempts at cladistic analysis were made by (as usual) Prof. Jacques Gauthier. Gauthier redefined the Archosauria as a crown group, the last common ancestor of birds and crocodiles and all of its descendants: crocs + rocs. Generally speaking, this nomenclature has stuck. Above Archosauria were the Erythrosuchidae and other forms previously referred to as "Proterosuchia." Gauthier used the term Archosauriformes to describe the clade uniting these forms with Archosauria.
A variety of names have been given to the two stem clades within Archosauria. Perhaps the most common are the Ornithodira (rocs > crocs) and Crurotarsi (crocs > rocs). The Ornithodira were originally the Ornithosuchia -- named after the Ornithosuchidae -- until it was learned that the Ornithosuchidae were more likely crurotarsans, which caused a certain amount of embarrassment all around. In any event, the Ornithodira are now believed to include Scleromochlus, pterosaurs, dinosaurs, and birds. Crurotarsi is believed to include crocs and all other "thecodonts" within the Archosauria: phytosaurs, aetosaurs, rauisuchians, ornithosuchids, and many strange and poorly-known beasts like Doswellia.
Unfortunately, the branching order within Crurotarsi has been, so far, impervious to cladistic analysis. Successive analyses by Benton & Clark (1988), Sereno (1991), Parrish (1993), Juul (1994), Benton (1999), and Benton & Walker (2002) have yielded rather discordant results, as shown in the table below (cladograms simplified and nomenclature standardized to promote comparison).
Archosauria
├─Ornithodira
│ ├─Ornithosuchidae
│ └─┬─Pterosauria
│ └─Dinosauria
└─Crurotarsi
├─Phytosauridae
└─┬─Gracilisuchus
└─┬─┬─Aetosauridae
│ └─Prestosuchidae
└─┬─Rauisuchidae
└─Crocodylomorpha
|
Archosauria
├─Ornithodira
│ ├─Pterosauria
│ └─Dinosauria
└─Crurotarsi
├─Phytosauridae
└─┬─Ornithosuchidae
└─┬─Aetosauridae
├─Gracilisuchus
├─Prestosuchidae
├─Rauisuchidae
└─Crocodylomorpha
|
Archosauria
├─Ornithodira
│ ├─Pterosauria
│ └─Dinosauria
└─Crurotarsi
├─Ornithosuchidae
└─┬─Phytosauridae
└─┬─Prestosuchidae
└─┬─Aetosauridae
└─┬─Rauisuchidae
└─┬─Gracilisuchus
└─Crocodylomorpha
|
|
Archosauria
├─Ornithodira
│ ├─Pterosauria
│ └─Dinosauria
└─Crurotarsi
├─┬─Prestosuchidae
│ └─Aetosauridae
├─Phytosauridae
└─┬─Ornithosuchidae
└─┬─┬─Gracilisuchus
│ └─Rauisuchidae
└─Crocodylomorpha
|
Archosauria
├─Ornithodira
│ ├─Pterosauria
│ └─Dinosauria
└─Crurotarsi
├─Phytosauridae
└─┬─Aetosauridae
└─┬─┬─Ornithosuchidae
│ └─Prestosuchidae
└─┬─Gracilisuchus
└─┬─Rauisuchidae
└─Crocodylomorpha
|
Archosauria
├─Ornithodira
│ ├─Pterosauria
│ └─Dinosauria
└─Crurotarsi
├─Phytosauridae
└─┬─┬─Ornithosuchidae
│ └─┬─Prestosuchidae
│ └─Rauisuchidae
└─┬─Aetosauridae
└─┬─Gracilisuchus
└─Crocodylomorpha
|
Each of these studies has its strengths and weaknesses. For example, Sereno's study may have too few characters to resolve the number of taxa in the study. Parrish seems to have too many ankle characters. Indeed, the "ankle phylogeny" has taken on a whole new life through this study. Parrish probably set out deliberately to test the ankle phylogenies by cladistic methods. At any rate his data is overwhelmingly weighted with ankle characters. Benton (1999) and Benton & Walker (2002) begin with Parrish's data matrix and add further characters. However, the effect of tarsal characters is still very strong. As Dyke (1998: 561) has noted: "[A]nkle characters do not contribute significantly to the broad shape of archosaur phylogeny. Indeed, their considered importance to the classification of the group is probably counter-productive."
 Gower (2002) has shown that braincase characters, used alone, generate a rather different pattern. Interestingly, when Gower's characters are combined with the Parrish - Benton data set, the resulting phylogeny may be much closer to Gower's result. This, in any case, was the basis of our "best guess" phylogeny. Further details are given with the cladogram at the end of this unit.
Gower (2002) has shown that braincase characters, used alone, generate a rather different pattern. Interestingly, when Gower's characters are combined with the Parrish - Benton data set, the resulting phylogeny may be much closer to Gower's result. This, in any case, was the basis of our "best guess" phylogeny. Further details are given with the cladogram at the end of this unit.
Finally, it has become de rigueur in reviews of archosaur phylogeny to add a few words about molecular phylogenies. Molecular phylogenies can add absolutely nothing to our understanding of basal archosaur evolution because all of the taxa of immediate interest are extinct. But, then again, who are we to depart from such a well-established scientific and literary tradition?
Accordingly, and merely by way of example, we include the enlightening cladogram of Hedges & Polling (1999):
These authors (with truly stunning arrogance) note: "determining how the many groups of extinct reptiles of the late Paleozoic and early Mesozoic fit into this molecular phylogeny will be a challenge to paleontologists."
Yes, indeed .... ATW040123.
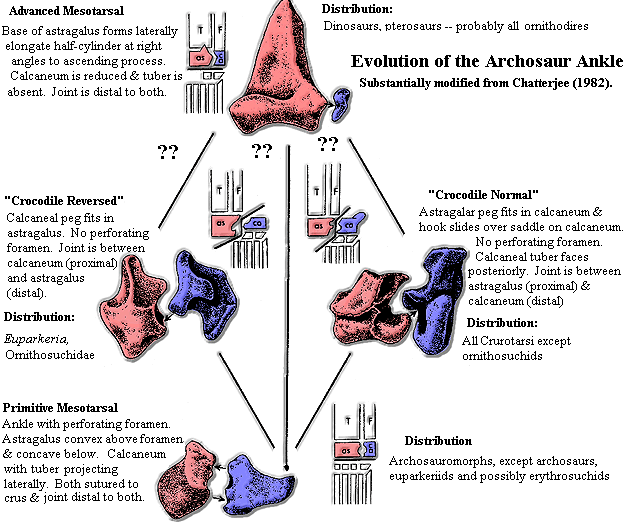 A Footnote on Ankles
A Footnote on AnklesConsidering the historical importance of the archosaur tarsus, we would be remiss if we failed to provide at least some information on the structures in question. The diagram at right was adapted from Chatterjee (1982) and has been somewhat modified from the original in light of later discoveries. It is more or less self-explanatory.
The basal archosauriform tarsus consisted of an astragalus and calcaneum sutured to the lower leg bones. the two proximal tarsals articulated by means of reciprocal peg and socket joints above and below a large perforating foramen. The functional ankle joint was between the proximal and distal tarsals, i.e. below the astragalus and calcaneum. The calcaneum bore a large, projecting tubercle which stuck straight out to the side.
Most Crurotarsi (the croc lineage of archosaurs), modify this arrangement. They have the "crocodile normal" tarsus. Here, the lower peg on the astragalus still articulates with a socket on the calcaneum. However, the upper articulation has been transformed into a sliding, rotary contact in which a "hook" on the astragalus rotates around a hemispherical saddle on the calcaneum. The ankle joint is thus between the astragalus, which is still sutured to the lower leg bones, and the calcaneum, which is now functionally part of the foot. The perforating foramen has disappeared, and the calcaneal tuber projects to the rear.
In theory, the "crocodile reversed" tarsus involves the converse arrangement, in which the calcaneum functions as part of the crus and the astragalus joins the foot. However, there is some uncertainty about the existence of this morph. The ankles of living ornithosuchids and Euparkeria may have had rather complex topologies with alternate functional modes.
The "advanced mesotarsal" joint is placed once again between the proximal tarsals and the metatarsals. Here, the calcaneum is reduced to a minor element. The astragalus is very large, with strong sutural connections to the lower leg bones reinforced with an ascending process which follows the shaft of the tibia. In Chatterjee's diagram, this hinge-like joint is shown as being derived from one or the other crurotarsal form. This need not have been the case. We know so little about the basal radiation of ornithodires that nearly anything is possible.
ATW040124.
Archosauria: birds + crocs.
from the Early Triassic
Archosauriformes::: Proterochampsidae + * : Crurotarsi + Ornithodira.
No parietal (pineal) foramen or fossa [P93]; foramina for internal carotids lateral to basipterygoid process [P93]; no palatal teeth on pterygoid, palatine or vomer [$S91] [$π03]; vertebral intercentra absent [$P93]; calcaneum with hemicylindrical facet for astragalus [P93]; calcaneal tuber with at least some posterior deviation from lateral projection [P93]; calcaneal tuber taller than broad [P93]; calcaneal tuber projects posteriorly at 90° to long axis of articulation between crus & proximal tarsals [$P93]; calcaneal tuber deflected >45° posterolaterally [$S91] [$π03]; calcaneal tuber flares such that it is laterally concave & medially convex [$P93]; calcaneal facet for fibula continuous with facet for distal tarsal IV [$S91] [P93] [$π03]; dermal armor present [P93].
Notes: "[π03]" refers to our own analysis of published data.
Links: link.
References: Parrish (1993) [P93], Sereno (1991) [S91]. ATW040117.
checked ATW050702
Using this material. All material by ATW is public domain and may be freely used in any way (also any material jointly written by ATW and MAK). All material by MAK is licensed Creative Commons Attribution License Version 3.0, and may be freely used provided acknowedgement is given. All Wikipedia material is either Gnu Open Source or Creative Commons (see original Wikipedia page for details). Other graphics are copyright their respective owners