Cynodontia: Basal Cynodonts: Epicynodontia
Taxa on This Page
- Cynosaurus X
- Epicynodontia
- Galesauridae X
- Galesaurus X
- Nanictosaurus X
- Platycraniellus X
- Thrinaxodon X
- Tribolodon X
The Epicynodonts: Early Triassic Cynodonts
 The Galesauridae and the very well-known Thrinaxodonmake up the basal epicynodonts. The epicynodonts are similar to, but more mammal-like than, their ancestors the Permian procynosuchids. Basal epicynodonts were small, solidly built carnivores, capable of running, to judge by the erect posture of their strong, if short, hind legs. An animal like Thrinaxodon, in its cranial anatomy and dental morphology must be close to the ancestry of the earliest mammals. Certain of its cheek teeth hear a striking similarity to those of the latest Triassic mammals.
The Galesauridae and the very well-known Thrinaxodonmake up the basal epicynodonts. The epicynodonts are similar to, but more mammal-like than, their ancestors the Permian procynosuchids. Basal epicynodonts were small, solidly built carnivores, capable of running, to judge by the erect posture of their strong, if short, hind legs. An animal like Thrinaxodon, in its cranial anatomy and dental morphology must be close to the ancestry of the earliest mammals. Certain of its cheek teeth hear a striking similarity to those of the latest Triassic mammals.
Another mammalian trend is seen in the lower jaw of Thrinaxodon. The teeth on either side were set into a single bone, the dentary, which had become larger at the expense of the smaller bones at the back of the jaw. The effect of this trend among cynodonts, toward a single lower jaw bone, was to make the jaws stronger. The zygomatic arch of bone at the rear of the skull, behind the eyes, was also was heavier; the masseteric fossa and coronoid process were essentially mammalian in size and shape, indicating the presence of well-differentiated masseter and temporal muscles. As with the procynosuchids, there is a complete, ossified, secondary palate, which completely covered the roof of the mouth and separated the breathing passage from the mouth. This development allowed the animal to breathe at the same time as retaining food in its mouth for longer periods, chewing it up into small pieces for quicker digestion.
The body was long and - for the first time among vertebrates - divided into a distinct chest (thoracic) and lower back (lumbar) region. The division was marked by the extent of the ribs; only the thoracic vertebrae bore ribs, and these formed a distinct cage, which housed the vital organs such as the heart and lungs. As in living mammals, there was probably a sheet of muscular tissue, the diaphragm, that closed off the rib cage in Thrinaxodon. As the animal breathed, the movement of the diaphragm would have filled and emptied its lungs efficiently an essential development in the evolution of body temperature control.
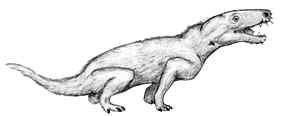 Many other structural changes had occurred showed a number of progressive changes over their Permian procynosuchid ancestors. For example, one of the foot bones had developed a heel which, with the help of strong tendons, would have acted as a lever to lift the foot clear of the ground with each step. The toes were all of equal length, allowing the body weight to be evenly distributed over them. MAK000718 ATW030212
Many other structural changes had occurred showed a number of progressive changes over their Permian procynosuchid ancestors. For example, one of the foot bones had developed a heel which, with the help of strong tendons, would have acted as a lever to lift the foot clear of the ground with each step. The toes were all of equal length, allowing the body weight to be evenly distributed over them. MAK000718 ATW030212
Image: Thrinaxodon (left) from drawing by Professor Paul E. Olsen - The Triassic World.
Descriptions
Epicynodontia: Galesaurus + Galileo
Range: from the Late Permian
Phylogeny: Cynodontia :: Procynosuchidae + * : Galesauridae + (Thrinaxodon + Eucynodontia)
fusion of parietals at midline [S01]; zygomatic more robust and broader than in basal forms [K82]; squamosal flares posteriorly in posterior wall of temporal fossa [HB86]; squamosal auditory meatus groove moderately deep (deeper than basal cynodonts) [SH98] [HK01$]; squamosal with enlarged descending flange lateral to quadratojugal [SH98] [HB86$]; quadrate & quadratojugal smaller and more closely integrated into squamosal [K82] [HB86$]; quadrate contacts only squamosal & not paroccipital process [SH98]; occiput wide & low [K82]; posttemporal foramen reduced [HB86]; paroccipital process expanded anteroposteriorly [HB86]; anterodorsal process of prootic extensively contacts epipterygoid [HB86]; secondary palate complete in most except for small anterior fissure [K82] (contra [H91]); ectopterygoid reduced [K82]; pterygoid lateral processes slight [K82]; interpterygoid vacuity closed in adults [K82] [HB86$] [HK01$]; dentary larger & post-dentary bones smaller than in procynosuchids [K82]; coronoid process very large & extends dorsally well into level of temporal fenestra [K82]; coronoid process dorsal limit above level of orbit [SH98] [HB86$] or to mid-orbit [HK01$]; masseteric fossa extends to ventral margin of dentary [SH98] [HB86$] [HK01$]; height of postdentary bones less than 50% dentary height [SH98]; intramandibular fenestra between surangular & angular covered by dentary [SH98]; 4 upper incisors [SH98] [HK01$]; incisors 4/3 [K82] [SH98] [HB86$] [HK01$]; precanines absent [K82] [HB86$]; 7-9 postdentary (sic -- postcanine?) teeth [K82]; postcanines with accessory cingular cusps enlarged mesial & distal to main cusp, with rest of cingulum reduced or absent [K82]; vertebrae amphicoelous [SH98]; ribs with costal plates [SH98] [H91] [HK01$]; posterior process of ilium longer than acetabulum diameter [SH98] [HK01$]; pubis length shorter than diameter of acetabulum [SH98] [HK01$]; pes 4 with 4 phalanges [SH98].
References: Carroll (1988); Hopson (1991) [H91], Hopson & Barghusen (1986) [HB86]; Hopson & Kitching 2001) [HK01]; Kemp (1982) [K82]; Sidor 2001) [S01]; Sidor & Hopson 1998) [SH98]. ATW030722.
Galesauridae:
Range: Late Permian to Middle Triassic of Africa, Europe, Antarctica, Russia & probably everywhere else.
Phylogeny: Epicynodontia : (Thrinaxodon + Eucynodontia) + * : Cynosaurus + Galesaurus + Nanictosaurus + Platycraniellus + Tribolodon.
Characters: snout broader than tall [SH98]; zygomatic arch enlarged; complete secondary palate; coronoid process with masseteric fossa; all postcanines on lower jaw on dentary; mesial accessory cusps absent [HB86$]; lower lingual cingulum absent [SH98] [HB86$]; distinct lumbar & thoracic regions, with ribs restricted to thoracic region; limbs somewhat more upright; calcaneal heel; diaphragm probably completely established.
References: Hopson & Barghusen (1986) [HB86]; Kemp (1982) [K82]; Sidor & Hopson (1998) [SH98]. ATW030215.
Cynosaurus: Genus Cynosaurus Schmidt 1927) C. suppostus (Owen 1876) (= Cynosuchus Owen 1876 = Cynosuchoides Broom 1931 = A'anictosaurus Broom 1936 = Mygalesuchus Broom 1942 = Baurocynodon Brink 1951).
Range: Late Permian of South Africa, Upper Daptocephalus Zone. [K82]
Phylogeny: Galesauridae : Galesaurus + Nanictosaurus + Platycraniellus + Tribolodon + *.
Characters: secondary palate not complete [K82]; teeth similar to Thrinaxodon, but without cingulum [K82].
References: Kemp (1982) [K82]. MAK 000718. ATW030419.
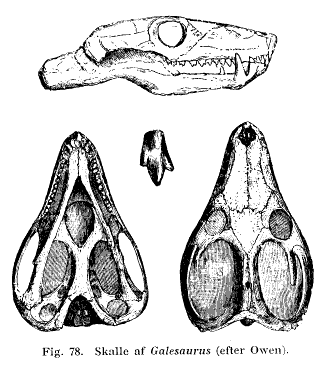 Galesaurus: (= Glochinodon van Hoepen 1916 = Glochinodontoides Haughton 1924b). G. planiceps Owen 1859 (= Glochinodon detinens van Hoepen 1916 = Glochinodontoides gracilis Ilaughton 1924b = Notictosaurus gracilis Broom & Robinson 1948a = Notictosaurus trigonocephalus Brink & Kitching
1951b).
Galesaurus: (= Glochinodon van Hoepen 1916 = Glochinodontoides Haughton 1924b). G. planiceps Owen 1859 (= Glochinodon detinens van Hoepen 1916 = Glochinodontoides gracilis Ilaughton 1924b = Notictosaurus gracilis Broom & Robinson 1948a = Notictosaurus trigonocephalus Brink & Kitching
1951b).
Range: Early Triassic of South Africa, Lystrosaurus Zone. (Possibly Permian)
Phylogeny: Galesauridae : Cynosaurus + Nanictosaurus + Platycraniellus + Tribolodon + *.
Characters: frontals with anterior "spine" projections inserting between nasals, persisting in adults; postcanines with large central cusp curving back to overhang distal accessory cusp [K82]; postcanines lack mesial cingula & mesial accessory cusps [K82].
Links: Triassic Dinosauria (historical interest); On the Classification of the Dinosauria (1870) (same); The Permo-Triassic Extinction; 68 Människan. Hennes uppkomst och utveckling) (Swedish?); The Permo-Triassic Extinction; therapsid8b;
Note: Small, immature specimens have usually been referred to the genus Galesaurus, large, mature specimens to Glochinodontoides. MAK 000718.
References: Kemp (1982) [K82]. ATW030220.
Nanictosaurus: N. kitchingi Broom 1936 (= N. robustus Broom 1940b = N. rubidgei Brink & Kitching 1953).
Range: Late Permian of South Africa, Upper Daptocephalus Zone, Dicynodon laterticeps - Whaitsia Assemblage Zone.
Phylogeny: Galesauridae : Cynosaurus + Galesaurus + Platycraniellus + Tribolodon + *.
Note: N. robustus was previously considered a Procynosuchid and synonym of Procynosuchus delaharpeae; it is now known to be the same as N. kitchingi. MAK 000718. ATW040103.
Platycraniellus: = Platycranion van Hoepen 1916 = Platycranium van Hoepen 1917) P. elegans vanHoepen 1916 (= Platycranion elegans van Hoepen 1916 = Platycraniellus elegans van Hoepen 1917).
Range: Early Triassic of South Africa, Lystrosaurus Zone.
Phylogeny: Galesauridae : Cynosaurus + Galesaurus + Nanictosaurus + Tribolodon + *.
Remarks: This species is known with certainty only from the type, which comes from Liarrismith, Orange Free State. A second specimen referred to this species by Brink is a Galesaurus. MAK 000718.
Tribolodon: Seeley 1894a. T. frerensis Seeley 1894a.
Range: Early Triassic of South Africa, Cynognathus Zone. [K82]
Phylogeny: Galesauridae : Cynosaurus + Galesaurus + Nanictosaurus + Platycraniellus + *.
Characters: postcanine teeth tall & slender, with strong accessory cusps mesial & distal [K82]; labial cingulum present with additional accessory cusps [K82].
Note: [1] The name appears to be preoccupied by TribolodonGünther 1877, a genus of cyprinid fish (dace). [2] This is the youngest species referable with certainty to the Galesauridae. Though frequently classified as a cynognathid, it is a typical galesaurid in its known features.
References: Kemp (1982) [K82]. ATW030215.
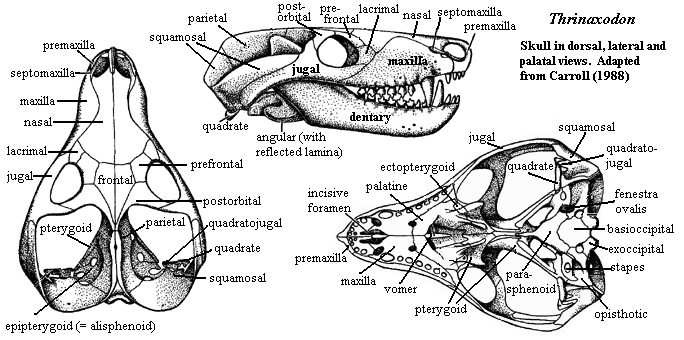 Thrinaxodon: T. liorhinus Seeley 1894a.
Thrinaxodon: T. liorhinus Seeley 1894a.
Range: Early Triassic Lystrosaurus Zone of South Africa & Fremouw Formation of Antarctica. [K82].
50 cm badger-like carnivore known from many remains [C88]; snout about as broad as tall [SH98]; postorbital and postfrontal merge [C88]; narrow braincase in sharp parietal crest [C88]; pineal opening present [C88]; robust zygomatic arch; adductor chamber almost half length of skull; squamosal covers posterior and forms most of adductor chamber & lateral surface of occiput [C88]; quadrate and quadratojugal much reduced & fit into sockets on squamosal [C88]; quadrate with posterior flange inserting in squamosal & capable of some anteroposterior motion [K82]; double condyle on occiput, but condyles remain below foramen magnum; ethmoturbinates present of type associated with homeothermic water-recovery system [H94]; fully ossified secondary palate [C88] [HB86]; incisive foramina present (shared with eucynodonts) [SH98]; palatine flanges in secondary palate meets at midline [SH98]; vomers with parallel sides, not expanded [SH98]; large dentary with coronoid reaching above zygomatic arch [C88]; masseteric fossa present and "reaches to its base" (base of dentary? or coronoid process?) [C88]; post-dentary bones lose sutural contact with dentary; reflected lamina large and deflected laterally [C88]; no evidence of a dentary- squamosal articulation [C88]; may have been a partial surangular - squamosal articulation [K82]; teeth with "reptilian" alternate replacement pattern [K82] [2]; dental formula 4/3,1/1,7-9/7-9 [C88]; cheek teeth regularly replaced, don't occlude, but have multiple cusps [C88]; upper cingulum much 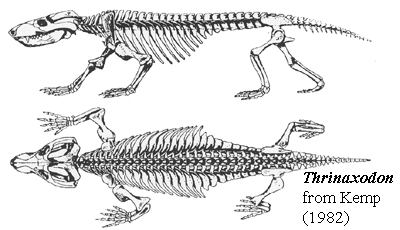 reduced or absent [K82]; lower lingual cingulum small [SH98]; proatlas present, but simplified atlas-axis articulation permits some rotation of head; long trunk; zygapophyses on dorsal vertebrae strongly angled upwards (limits lateral, but permits dorsoventral flexion) [C88]; zygapophyses with accessory articulations between peg below postzygapophyses and groove below prezygapophysis [K82]; 7 cervicals,13 dorsals, 7 lumbar, 5 sacrals, 10-15 caudals [C88]; rib capitulum articulates with parapophysis formed equally by 2 vertebrae [K82]; ribs proximally expanded to form overlapping costal plates (lateral stiffening) [C88] [K82]; lumbar ribs have costal plates only [C88]; lumbar plates are vertical [K82]; tail short [C88]; limbs short [C88]; pectoral girdle remains primitive; ventral process of clavicle & interclavicle retained (resist lateral forces of sprawling posture) [C88]; coracoid somewhat reduced; scapula major weight-bearing element, directed vertically; scapular blade deeply concave, with narrowed base [K82]; coracoid plate reduced [K82]; small procoracoid present; dorsal (scapular) portion of glenoid almost flat, with restricting movement [K82]; humerus short, heavy, complex and horizontal [C88]; phalangeal count 23443 [C88]; pubes & ischium reduced [C88]; ilium expanded anteriorly [K82]; femur angled 55° outward, but capable of dorsoventral movement on angled humeral head; beginnings of gluteal-type musculature from iliofemoralis; greater trochanter moderately developed [K82]; calcaneal heel; simple hinge between astragalus (integrated with tibia) and calcaneum; gastrocnemius attaches to calcaneal tuber; plantigrade stance [C88].
reduced or absent [K82]; lower lingual cingulum small [SH98]; proatlas present, but simplified atlas-axis articulation permits some rotation of head; long trunk; zygapophyses on dorsal vertebrae strongly angled upwards (limits lateral, but permits dorsoventral flexion) [C88]; zygapophyses with accessory articulations between peg below postzygapophyses and groove below prezygapophysis [K82]; 7 cervicals,13 dorsals, 7 lumbar, 5 sacrals, 10-15 caudals [C88]; rib capitulum articulates with parapophysis formed equally by 2 vertebrae [K82]; ribs proximally expanded to form overlapping costal plates (lateral stiffening) [C88] [K82]; lumbar ribs have costal plates only [C88]; lumbar plates are vertical [K82]; tail short [C88]; limbs short [C88]; pectoral girdle remains primitive; ventral process of clavicle & interclavicle retained (resist lateral forces of sprawling posture) [C88]; coracoid somewhat reduced; scapula major weight-bearing element, directed vertically; scapular blade deeply concave, with narrowed base [K82]; coracoid plate reduced [K82]; small procoracoid present; dorsal (scapular) portion of glenoid almost flat, with restricting movement [K82]; humerus short, heavy, complex and horizontal [C88]; phalangeal count 23443 [C88]; pubes & ischium reduced [C88]; ilium expanded anteriorly [K82]; femur angled 55° outward, but capable of dorsoventral movement on angled humeral head; beginnings of gluteal-type musculature from iliofemoralis; greater trochanter moderately developed [K82]; calcaneal heel; simple hinge between astragalus (integrated with tibia) and calcaneum; gastrocnemius attaches to calcaneal tuber; plantigrade stance [C88].
Notes: [1] The best known epicynodont, and along with Cynognathus the best known cynodont. Nythosaurus larvatus Owen is based on the natural mould of a skull bearing impressions of the postcanine teeth. It is probably synonymous with either Thrinaxodon liorhinus or Platycraniellus elegans. MAK 000718. [2] in the typical sauropsid pattern, waves of tooth replacement move down each ramus of the jaw, keeping more or less in synchrony. The replacement wave affects only alternate teeth, avoiding any long gaps. The next replacement wave affects the teeth left intact on the first wave. The pattern in Thrinaxodon was somewhat more complex (i.e. we didn't understand Kemp's explanation), but followed the same general scheme.
Links: Thrinaxodon CAT Scan Rendering; Thrinaxodon CAT Scan Rendering; Estes, R., 1961; Thrinaxodon; TMM - Exhibits - Scratching the Surface - Thrinaxodon; Thrinaxodon sp.; Later Mammal-like Reptiles; Lecture
9 - Triassic: Moenkopi, Karroo, Ischigualasto; Ungewöhnliche Krokodilkonstruktionen der Vorzeit:; Mammalogy
- Lecture 3; ?????????? a great antidote to Bakker's unbearably cute image).
References: Carroll (1988) [C88]; Hillenius (1994) [H94]; Kemp (1982) [K82]; Sidor & Hopson (1998) [SH98]. ATW030212.

