 Dinilysia
Dinilysia| Squamata | ||
| The Vertebrates | Pythonomorpha: Alethinophidia |
| Vertebrates Home | Vertebrate | Vertebrate |
|
Abbreviated Dendrogram
Squamata
├─Iguania
└─Scleroglossa
│
└─Pythonomorpha
├─Mosasauroidea
└─Serpentes
├─Scolecophidia
└─Alethinophidia
├─Dinilysia
└─┬─Aniloidea
│ ├─Aniliidae
│ └─┬─Cylindrophiidae
│ └─┬─Uropeltidae
│ └─Anomochilidae
└─Macrostomata
├─Boidae
└─Caenophidia
├─Acrochordoidea
└─Colubroidea
├─Colubridae
└─Viperidae
|
Contents
Overview |
Serpentes: Scolecophidia + * : Dinilysia + (Aniloidea + Macrostomata).
vomerine process of premaxilla large and broadly in contact with vomers; $ vomeronasal nerve enters vomer through single large (or 1 large + 1 small) foramen [T[; coronoid present; maxilla not mobile; ascending process of maxilla contacts frontal; rostral region articulated to braincase by naso-frontal articulation; braincase narrow, elongate and tapers anteriorly; postfrontal well-developed and fused to parietal; quadrate short & vertical; footplate of stapes large; facets of zygapophyses slant upward; middle and posterior vertebrae lack hypapophyses.
Note: these are the primitive conditions, i.e. characteristics of the paraphyletic Aniloidea.
Links:Lab VII. Snakes a sampler); S0952836999005051d 49..58 taxonomy, feeding biology and evolution); Digimorph - Ramphotyphlops braminus (Brahminy blind snake) - head; Herpetology- Snakes; PII- S1631-0691(02)01510-X.
References: Holman 2000); Tchernov et al. 2000) [T] ATW030923
Range: upK (Coniacian [C&A]) of SAm.
Phylogeny: Alethinophoidia :(Aniloidea + Macrostomata) + *.
Characters: slightly flattened skull [C&A]; palatine does not contact ectopterygoid; pterygoid with teeth; outer orbital (lateral) margin of prefrontal vertical; low medial foot process of prefrontal; supraorbital present(?); quadrate vertical; shaft of stapes not straight; inhabited seasonally (or sporadically?) arid interdune environment with wind-dominated sedimentation [C&A]; terrestrial, semi-fossorial mode of life, with active or ambush predation in loose groundcover [C&A].
Links: BBC Online - Walking with Dinosaurs - Fact Files; Mike Caldwell hunts fossil snakes in Argentina.;
References: Caldwell & Albino (2001) [C&A]; Tchernov et al. (2000) [T]
Note: shares almost no derived features with scolecophidians.
Aniloidea: Anilius > Crotalus.
Range: fr upK of NAm, Eur, SAm, India & SW Asia.
Phylogeny: Alethinophidia ::Macrostomata + * : Aniliidae + (Cylindrophiidae + Uropeltidae + Anomochilidae)). 010901.
 Aniliidae:Anilius, Coniophis, Eoanilius (?).
Aniliidae:Anilius, Coniophis, Eoanilius (?).
from the Late Cretaceous (Campanian). Currently SAm only (1 sp.)
Aniloidea :(Cylindrophiidae + (Uropeltidae + Anomochilidae)) + *.
Robust, tightly knit skull; premaxillary, palatine, and pterygoid teeth present; lower jaw hinged on surangular; palatine has gliding joint on maxilla; m. retractor vomeris present; splenial absent; gape not wide; upper jaw solidly attached to skull; frontal-parietal articulation closed & immobile; reduced eyes lying beneath large, at least semi-transparent head scales; squamosal present, but short; supratemporal present; fovea dentis present in occipital condyle; vertebrae squarish in outline and not longer than broad; zygosphenes flare upward; vertebrae lack prominent neural spine or substantial accessory processes; hemal carina absent; hypapophyses absent on mid-thoracic vertebrae; vestigial hindlimbs and girdle; single bone in pelvic girdle; femur may project as anal spur; small, smooth scales; ventral scales slightly larger than dorsal; ventral scales with black & white checkered pattern; gastrosteges present; liver relatively anterior; heart relatively anterior; 2 lungs, with left lung reduced; kidneys relatively anterior; fossorial; carnivorous; ovoviviparous.
Note: At this level, snake phylogeny is very unclear. Possibly the majority opinion is that the advanced snakes are more closely related to Boids than to *, although both groups are fairly primitive. Most aniliids use defensive display behaviors which attempt to mimic toxic or dangerous organisms. This group may also include Loxocemids. 010609.
Discussion:Aniliids today are represented only by the genus Anilius of South America. However, they are perhaps the oldest group of snakes known from reasonably distinctive fossil remains. They have a more or less continuous fossil record from the Upper Cretaceous to the present day.
The distinctive squarish vertebrae of Coniophis are known from numerous locations in the Upper Cretaceous, Paleocene and Eocene of North America. Hecht (1959); Holman (2000). Oddly, Coniophis is also known from the Cretaceous of South America -- one of several indications that a land bridge existed between the two continents at one or more times in the later Cretaceous. Holman (2000). The early Eocene was apparantly a good time for snakes in North America. However, the diversification seems to have been due to the evolution of larger, terresestrial forms. The fossorial aniliids are represented only by the familiar Coniophis. In the later Eocene, perhaps as the result of colder climates, many snake species became extinct, including Coniophis. Id.
Presumably, some relict population survived in South America, since Anilius, the False Coral Snake, is known from a number of locations in northern and Western South America today. However, with the exception of a few Miocene vertebrae (Colombophis) there does not appear to be any fossil record in the Western Hemisphere after the middle Eocene. It has been argued that Columbophis is more likely to be a uropeltid than a true aniliid. Szyndlar (1994). Given that Recent uropeltids are only found in a restricted area of South Asia, it is perhaps more likely yet that Columbophis represents some completely extinct branch derived from the the Coniophis stem.
About the same time as Coniophis was dying out in North America, Eoanilius was making its first appearance in Europe. Eoanilius is said to be a true aniliid and is a fairly common fossil snake (if any ophidian fossil can be referred to as "common") from the Eocene and Oligocene of England, France and Germany. Europe, like North America, suffered a devastating extinction of snake species during the later Eocene. However, through chance or some difference in circumstances, the aniliids appear to have hung on in Europe for at least some period of time. Szyndlar (1994). In fact, a possible aniliid, Michauxophis, has been reported from as late as the Pliocene of France. Bailon (1988).
Unfortunately, as with most areas of snake taxonomy, it is not clear that all workers are speaking about the same thing. The traditional practice was to lump the Cylindrophiidae, Aniliidae, Anomalochilidae, and Uropeltidae under a general heading: usually "Aniloidea," but also frequently "Aniliidae." There is certainly a distinct possibility that the Aniloidea, in this sense, are a clade which diverged early from the boids and their progeny. Cundall et al (1993) make a good case that any "aniloid" group would be paraphyletic. However, their cladistic analysis rests on the combination of skeletal and soft tissue traits. As they are careful to point out, their skeletal data, considered alone, would support an aniloid clade. Their soft tissue data, taken only from highly derived extant species, makes almost no sense at all if considered alone. It is only the combination of skeletal and non-skeletal characters which yields the paraphyletic aniloidea as used in these Notes. However, the combined tree is fairly robust, especially for a snake phylogeny. Furthermore, the supposition that all "aniloids" form a separate branch of the family tree requires us to accept an odd, mosaic distribution of traits.
In the last analysis, the apparant similarity of the aniloids is quite possibly due to the fact that all known members of the constituent groups are fossorial to one degree or another. The difficult business of crawling around underground creates some rather severe constraints on biological design. As touched on elsewhere, the pressures of becoming fossorial followed by regaining terrestriality, are likely to result in a radically re-engineered skull. However, the re-engineering is not possible without the initial step of fossorial existence. Thus the unique and radical kinesis of the advanced snakes sets them apart from the "aniloids," but also argues that advanced snakes descended from a fossorial stem group, presumably part of the original aniloid radiation. ATW 010108.
Note added in disproof: The Rieppel people, e.g. Tchernov et al. (2000), disagree on almost everything in Lee's trees, e.g. Scanlon & Lee (2000) -- everything except the monophyly of the aniloids. On this subject, they speak with one voice. Accordingly, I grudgingly concede error, with much whining and snarling, and have accordingly restored the Aniloidea to their presumably rightful place.
Links: Aniliidae; Animal Diversity Web: Anilius scytale: Media; Lab VII. Snakes; espacio.ya.com - Bienvenido a la página de aligatorjazz; s102110100043.pdf.
References: Bailon 1988); Cundall et al. (1993); Hecht (1959); Holman 2000); Scanlon & Lee (2000); Szyndlar (1994); Tchernov et al. (2000). ATW 010901.
 Cylindrophiidae(= Cylindrophiinae): Coniophis (?), Cylindrophis.
Cylindrophiidae(= Cylindrophiinae): Coniophis (?), Cylindrophis.
Recent of SE Asia.
Phylogeny: Aniloidea ::(Uropeltidae + Anomochilidae) + *.
Small to moderate size; teeth present on palatine & pterygoid; 9-13 maxillary teeth; robust dentary; anterior tips of dentary rounded, with limited, hinge-like mobility [LBC]; prominent mental groove; premaxillary process long, extending between nasals; large ectopterygoid, overlapping maxilla and pterygoid; prefrontal small; spectacle covering eyes present; postorbital present; frontal-parietal articulation closed & immobile; occipital condyle without fovea dentis; vertebrae generally same as in Aniliidae; heart ~30% of snout-to-vent length from snout; vestigial left lung complex present; kidneys relatively far posterior & near vent; scales smooth, with 19-23 rows; semi-aquatic. Many details of superficial characters (e.g. scales & dentition) at Cylindrophis maculatus.
Note: Eoanilius (upEc of Eur) has been referred to this taxon, but the assignment is strongly disputed.
Image: Cylindrophis rufus from foto-bibliotheek.
Links: Cylindrophiidae; Cylindrophis sp; Lady Di Body Art (Dutch); Cylindrophis maculatus (Best on the Web); Cylindrophis rufus (Japanese); Herpetofauna of Myanmar several images); 两栖动物分类 Chinese).
References: Cundall et al (1993); Cundall & Rossman (1993); Hecht (1959); Lee et al. (1999a) [LBC]. ATW040114.
 Uropeltidae
(= Uropeltinae): "shield-tail" snakes.
Uropeltidae
(= Uropeltinae): "shield-tail" snakes.
R of S Asia.
Phylogeny:Aniloidea :::Anomochilidae + *.
Skull massive and akinetic; usually pointed snout; upper jaw solidly attached to skull; premaxilla and maxilla closely associated sutured?); teeth absent from palatine & pterygoid; prefrontal extends between nasal and maxilla and interlocks strongly with maxilla; frontal-parietal articulation partially open; no eye-covering spectacles; small eyes; often have long posterior process on dorsal end of quadrate; quadrate associated directly with otic elements; supratemporal absent; basisphenoid, basioccipitals, prootics, and supraoccipitals all fused; inner centrum of atlas absent; long occipital condyle forms ball and socket joint with axis; fovea dentis present; no hind limb vestiges, but pelvic bone present; keratinous disk-shaped shield at tip of tail which may be specialized for prehensile or mimetic purposes; upper surface of tail with single or series of thickened scutes, often covered with spines; gastrosteges present but reduced, left lung very small; superficial branch of adductor mandibulae externalis originates from temporal tendon and inserts on coronoid process; heart located 30% of snout-to-vent length, liver about 50%; gall bladder near posterior limit of liver; kidney relatively far anterior to vent; carnivorous; fossorial; viviparous (?); mimetic defensive displays.
Links: Docent Webpage; Uropeltidae; espacio.ya.com - Bienvenido a la página de aligatorjazz (Spanish).
References: Cundall et al (1993); Cundall & Rossman (1993).
Image: Uropeltis from foto-bibliotheek. ATW020123.
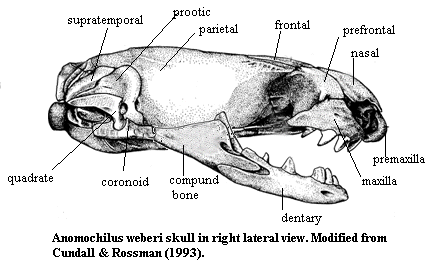 Anomochilidae: Anomochilus.
Anomochilidae: Anomochilus.
R of SE Asia.
Phylogeny:Aniloidea :::Uropeltidae + *.
Small, cylindrical body; head no wider than body; broad, blunt snout; pterygoid, palatine, pterygoid, and premaxillary teeth absent; 3-4 maxillary teeth; teeth short, with stout base; teeth not ankylosed & attachment may be fibrous; robust dentary; ectopterygoid much reduced, entirely incorporated in ligament between pterygoid and maxilla, but does not contact maxilla or pterygoid; mental groove absent; coronoid present & well-developed; nasal process of premaxilla weak and not fixed to nasals; nasals very large and participate in anterior margin of rostrum; prefrontal extends between nasal & maxilla and is supported on maxilla; frontal-parietal articulation partially open; process of parietal ("postfrontal") excludes frontal from margin of orbit; braincase relatively wide, lacking prominent crests or laterally expanded otic capsule; long posterior process on dorsal end of quadrate; separate supra-, ex- & basioccipitals and prootic; occipital condyle with fovea dentis; superficial adductor mandibulae externis originates on braincase only and inserts on coronoid process (?); spectacle scales reduced or absent; eyes reduced; smooth scales with 17 - 19 rows; rectal cecum absent; liver large and posteriorly asymmetrical; gallbladder displaced posteriorly; left lung complex absent; heart and lung relatively far anterior; 2 renal arteries; oviparous.
Links: Anomochilidae.
References: Cundall et al (1993); Cundall & Rossman (1993). 010107.
checked ATW060222
{kind=link}