The Berriasian Age
The Berriasian Age of Neocomian Time: 146 to 140 million years ago
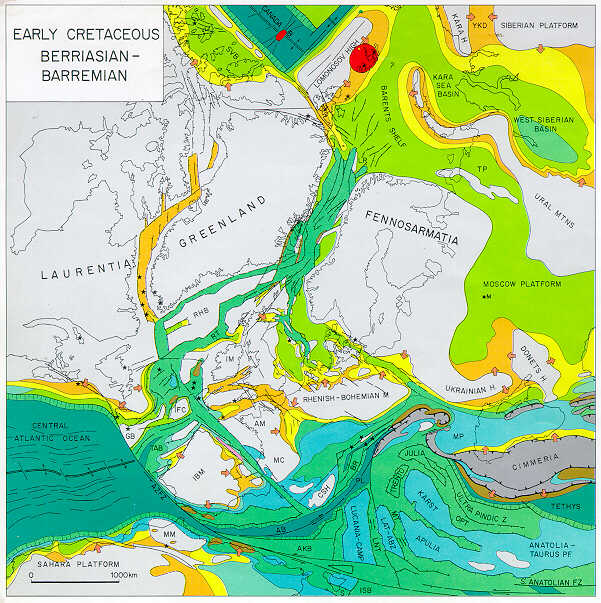
 The Berriasian is the first age of the Cretaceous. By the Berriasian, East Gondwana (India, Australia and Antarctica) had separated cleanly from West Gondwana (Africa and South America); and, by the end of the Age, India was beginning to break from East Gondwana. The Tethys had long since split the Gondwanan lands from the Northern lands, although the situation in the Caribbean Basin was complex. The Pacific Plate was growing, but still quite small, and the entire western margin of North and South America was fringed with volcanic island arcs. Zharkov et al. (1998).
The Berriasian is the first age of the Cretaceous. By the Berriasian, East Gondwana (India, Australia and Antarctica) had separated cleanly from West Gondwana (Africa and South America); and, by the end of the Age, India was beginning to break from East Gondwana. The Tethys had long since split the Gondwanan lands from the Northern lands, although the situation in the Caribbean Basin was complex. The Pacific Plate was growing, but still quite small, and the entire western margin of North and South America was fringed with volcanic island arcs. Zharkov et al. (1998).
 The cooling trend of the Late Jurassic continued into at least the middle of the Berriasian, and the poles may have borne small permanent ice caps, with glaciers at higher elevation in East Gondwana. The Atlantic and Ural connections between the Tethys and the northern sea were still well in the future, so that the Middle latitudes of the Northern hemisphere were thermally isolated and quite cool, at least by Mesozoic standards. Chumakov 2003).
The cooling trend of the Late Jurassic continued into at least the middle of the Berriasian, and the poles may have borne small permanent ice caps, with glaciers at higher elevation in East Gondwana. The Atlantic and Ural connections between the Tethys and the northern sea were still well in the future, so that the Middle latitudes of the Northern hemisphere were thermally isolated and quite cool, at least by Mesozoic standards. Chumakov 2003).
Most of the characteristic life forms of the Cretaceous had not yet evolved or were insignificant, including angiosperms, mosasaurs, therian mammals, hadrosaurs, rudists, calcareous plankton and planktonic foraminifera. Dinosaurs were represented by Brachiosaurus, Camptosaurus, Iguanodon, and a number of weird and poorly known transitional forms such as Echinodon, Embasaurus, Nuthetesand Stenopelix. Teleost fish were present, but not yet in their Cenozoic forms, and the dominant marine reptiles were Cimoliasaurus and Cryptocleidus.
Image credits: Camptosaurus from the College of Eastern Utah Museum site. Map from the American Association of Petroleum Geologists site.
Links: The Plesiosaur Site - Stratigraphy; DinoData Dinosaurs Timeperiods Berriasian; anrepeap.
ATW050219. Text public domain. No rights reserved.
Studies of the Berriasian and the Jurassic-Cretaceous boundary interval have been concentrated, particularly, in the area of the great former ocean of Tethys. This was the largest single geographical unit of the time and its rocks and fossils give the best possibilities for wide and consistent correlations. In the earliest Cretaceous, this gigantic ocean divided Laurasia from Gondwana, and Berriasian marine sediments are widespread from Mexico to the Russian Far East and Japan via north Africa, Europe, Crimea, Arabia, Iraq, Iran and Tibet.
Early research on the Berriasian was, in fact, not in Tethys nor even on marine sedimentary rocks, but on non-marine strata. The English Purbeck Formation is largely of Berriasian age, though it extends down into the Tithonian (Portlandian) and up into the Valanginian. And it was the Purbeck Formation and its rich molluscan and vertebrate fauna (mammals, lizards, pterosaurs, crocodilians etc) that took the attention of famous geologists such as De la Beche, Buckland, Webster and Fitton. Brongniart, in 1829, labelled these beds as part of the Jurassic, in the "Purbeckien" stage, a name officially suppressed in the 1960s: whereas D'Orbigny relegated the "Purbeckian" to the Cretaceous.
When geologists started to study marine sequences of this age in Tethys, the focus moved for a time, particularly, to the limestones of southern France, and a new name was introduced, Berriasian (after the village of Berrias in the Ardèche region: Coquand, 1869). As stratigraphers now view things, the Berriasian defines the lowest Cretaceous and its base delimits the Tithonian, the top of the Jurassic. Unfortunately, the section at Berrias has no macrofossils near where the boundary might be expected, and so specialists have been busy looking for better, more fossiliferous, sections that show the uppermost Tithonian and lowest Berriasian.
Historically, in the Mediterranean parts of Tethys, the Berriasian has been described as consisting of athe following ammonite zones and subzones:
| Fauriella boissieri Zone |
Thurmanniceras otopeta |
| Tirnovella alpillensis |
| Berriasella picteti |
| Malbosiceras paramimounum |
| Tirnovella occitanica Zone |
Dalmasiceras dalmasi |
| Berriasella privasensis |
| Tirnovella subalpina |
| Pseudosubplanites grandis Zone |
Pseudosubplanites grandis |
| Berriasella jacobi |
At two international meetings on the Cretaceous, stratigraphers considered the best level to place the base of the Berriasian, and formally voted that the base of the grandis and jacobi subzones afforded the optimum levels for this. Unfortunately, some of the best sections for the J/K boundary have few or no ammonites in the critical interval. Therefore, in the last thirty years, micropaleontology has become more and more important, as microfossils are more consistently present and much more common than ammonites in many profiles in Tethys. To the precision of microfossil ranges has been added another stratigraphic tool, magnetostratigraphy, which is being applied to key sections.
The latest Jurassic and earliest Cretaceous rocks are complicated and hard to decipher outside Tethys, as J/K palaeogeography is quite complicated. The more extensive late Jurassic seas had withdrawn from large areas and earlier seaways had closed. Some regions were periodically connected with Tethys, but others were isolated, and some that had been sea had become freshwater lakes and land, as in England, Germany and Poland with the Purbeck beds, which only have rare levels with marine fossils. Non-marine rocks of this age are also well-developed in Mongolia and China. Some areas nearer to the paleo-poles had limited endemic biotas (Siberia, Antarctica, Canada). Others had oceanic connections that sometimes allowed faunal migration, but they were very distant from Tethys (e.g. Argentina) and had their own particular marine faunas Some were landlocked basins with low biotic diversity (Russian Platform), and others have suffered subsequent extreme tectonic disruption (Japan). The challenge and task with all these areas is to correlate with the better documented sequences in Tethys.
A global reference profile for the base of the Berriasian (a GSSP) is under active consideration by the International Subcommission on Cretaceous Stratigraphy (ISCS) of IUGS, and a range of contender GSSP localities are currently being studied in detail by the ISCS's Berriasian Working Group.
Several markers are being employed to refine correlations and to work towards definition of a widely useable base for the Berriasian Stage. These include, particularly, calpionellids, calcareous nannofossils, such as Nannoconus, ammonites, palynological datums and magnetostratigraphy. Depending on which final fossil markers are employed to define a boundary , in magnetostratigraphic terms, it is likely to be between the base and top of magnetozone M19n. The calibration of markers, such as Nannoconus steinmannii minor, N. kamptneri minor, and Calpionella alpina, with precisely fixed magnetozones gives greater precision in trying to identify the best position for a boundary.
Bill Wimbledon 130211
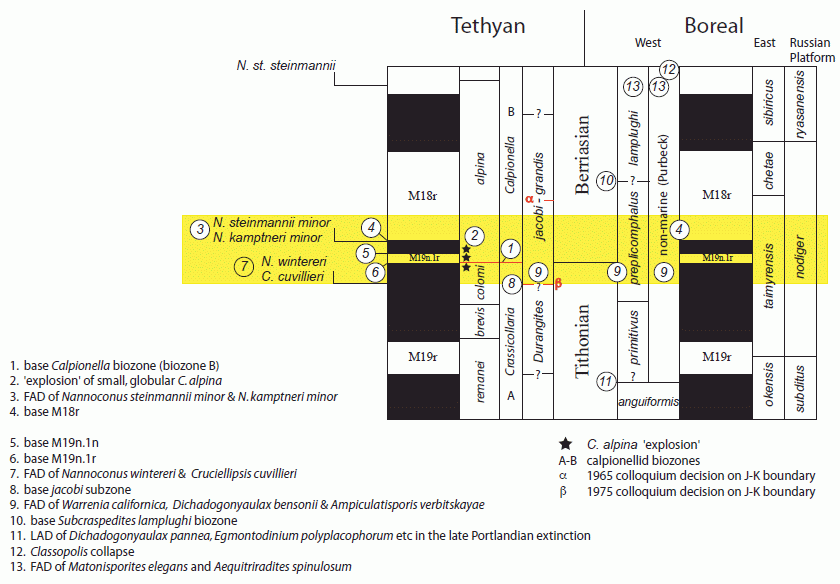
Markers useful in defining the Jurassic-Cretaceous Interval (yellow band). Adapted from Wimbledon et al 2011 p.299, fig.1
Berriasian Life
 Generally speaking, the Cretaceous was not a good time for arthropods (although it is not easy to be certain, because of the usually miserable fossil record). The clear exception are the Crustacea who produced both lobsters and shrimp at some point in the earliest Cretaceous. Sadly, such essential condiments as butter, tomato sauce, and horseradish were not yet available in the Berriasian.
Generally speaking, the Cretaceous was not a good time for arthropods (although it is not easy to be certain, because of the usually miserable fossil record). The clear exception are the Crustacea who produced both lobsters and shrimp at some point in the earliest Cretaceous. Sadly, such essential condiments as butter, tomato sauce, and horseradish were not yet available in the Berriasian.
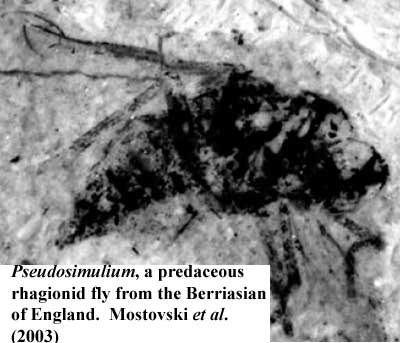
Chelicerates nearly became extinct in the Cretaceous, at least by comparison to their large diversity in the Paleozoic and Cenozoic. However, spiders were present and the remains of a number of orb-web weavers are known from the Berriasian. These spiders may have benefited from the large number of flying insects. The flying insect jobs were dominated by Diptera. In fact, it was an age of flies. Since many modern neopteran insect groups had not yet become important, flies occupied a large number of niches in which they are now marginal, including the job of nipping at vertebrates (absent gnats, midges and mosquitoes). Mostovski et al. (2003). When angiosperms arrived later in the Neocomian, it was these flies with suctorial mouthparts which first evolved as pollinators.
Two important clades of insect recyclers are first found in the Berriasian. The first of these are the Scarabaeoidea, the stag, scarab and dung beetles. They are probably of Jurassic origin, but diversified in the Berriasian. Krell 2000). A far more important development was the evolution of termites Isoptera), which evolved from the ancient cockroach lineage (Blattaria) during, or just before, the Berriasian. Mayhew 2002).
 Bear in mind that wood recycling had been handled almost exclusively by fungi since the Mississippian. The fungi do a very thorough job, but they're slow. Lignin is a fiendishly complex molecule, and generates an amusing variety of toxic phenols if it isn't digested in just the right way. Termites can't actually digest lignin themselves, but the termite gut is a very complex ecosystem which plays host to an entire battery of bacterial and protist symbiotes which cooperate to get the job done rather quickly. C.f. Oxymonadida one of the main protist groups involved in this industry). The advent of termites would likely have changed the entire forest food chain. Prior to termites, dead or fallen trees would have remained in place for many years as they were slowly consumed by rot. Termites recycle the nutrient content of wood much faster and convert it into insect biomass. This would not only permit faster recovery of woodlands from storm, fire and (perhaps) sauropod damage, but would open up vast new opportunities for small insectivores. One might suspect that the rapid spread of the opportunistic and aggressive angiosperms and the diversification of insectivorous therian mammals just after the Berriasian was not a coincidence. Thus, we may have mutant cockroaches to thank for the existence of both ourselves and our main food supplies.
Bear in mind that wood recycling had been handled almost exclusively by fungi since the Mississippian. The fungi do a very thorough job, but they're slow. Lignin is a fiendishly complex molecule, and generates an amusing variety of toxic phenols if it isn't digested in just the right way. Termites can't actually digest lignin themselves, but the termite gut is a very complex ecosystem which plays host to an entire battery of bacterial and protist symbiotes which cooperate to get the job done rather quickly. C.f. Oxymonadida one of the main protist groups involved in this industry). The advent of termites would likely have changed the entire forest food chain. Prior to termites, dead or fallen trees would have remained in place for many years as they were slowly consumed by rot. Termites recycle the nutrient content of wood much faster and convert it into insect biomass. This would not only permit faster recovery of woodlands from storm, fire and (perhaps) sauropod damage, but would open up vast new opportunities for small insectivores. One might suspect that the rapid spread of the opportunistic and aggressive angiosperms and the diversification of insectivorous therian mammals just after the Berriasian was not a coincidence. Thus, we may have mutant cockroaches to thank for the existence of both ourselves and our main food supplies.
Image Credits: termite from Fossilmall.com. Pseudosimilium from Mostovski et al. (2003).
ATW050220. Text public domain. No rights reserved.
ATW041026, MAK050104, Bill Wimbledon 130211. Material by ATW public domain; all other original material
Creative Commons Attribution
checked ATW060117