 Lepidosauriformes: Paliguana, Saurosternon. Lizards > plesiosaurs.
Lepidosauriformes: Paliguana, Saurosternon. Lizards > plesiosaurs.| Lepidosauromorpha | ||
| The Vertebrates | Lepidosauriformes |
| Vertebrates Home | Vertebrate | Vertebrate |
|
Abbreviated Dendrogram
○Diapsida
├─Archosauromorpha
└─○Lepidosauromorpha
├─Sauropterygia
└─○Lepidosauriformes
├─Kuehneosauridae
└─┬─Marmoretta
└─┬─Sophineta
└─○Lepidosauria
├─Sphenodontia
└─Squamata
|
Contents
Overview |
Some years ago, Paliguanaheld a position of some prominence at this point in these phylogenetic proceedings. Simultaneously with the posting of this essay, the Paliguanidae disappeared from the site, just as they disappeared from the South African Karoo in the Early Triassic, 240 million years ago. Fortunately, this second mass extinction is somewhat less enigmaticand may only be temporary. However, it is an appropriate occasion to say a few words about the use and misuse of stem taxa.
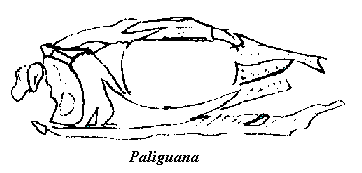
Paliguana is known only from a partial skull described by Broome in 1903. It was immediately recognized as very lizard-like, as was (Huxley's?) Paleagama (or Palæagama), which is also known from a single, fairly complete, but badly preserved, specimen. Somewhat later, Saurosternon, a headless body with similar characteristics, was also recovered from similar exposures. Clearly, these were all basal diapsids -- too primitive to be lizards, but fairly close morphologically to that line and to each other. The temptation to put the ambiguous body of Paleagama together with the isolated skull of Paliguana proved too much, and so the Paliguanidae were born, the gaps being filled in with material from Saurosternon. In truth, they are a good match and, as Carroll 1988a) has pointed out, there is nothing which prevents them from belonging to the same, or closely related, genera.
However, the advent of computer cladistics had curious effects on the Paliguanidae. The first significant cladistic analysis of the basal diapsids was performed by Gauthier et al. 1988) and published back-to-back with Carroll's (1988a) defense of the Paliguanidae. The Gauthier group simultaneously conceived the taxon Lepidosauriformes. Their cladogram looked like this:
Some of their analysis was attacked by Carroll (1988a) and others -- not without reason, as we shall see. After several years of good-natured academic banter, Clark & Hernandez 1994) published a description of an interesting new amphisbaenid-like creature, Tamaulipasaurus. In the process of discussing the phylogenetic relationships of this interesting new find, the authors off-handedly did a remarkable job of synthesizing the criticisms of the original cladistic study. Further, they did not stop at carping, but eliminated the offending characters, added new species, rescored the errors, and reran the analysis with a slightly more sophisticated software package. In a noble attempt to avert battles over mere nomenclature, these authors used numbers, rather than names, for the nodes. Their result, somewhat abbreviated, looks like this:
So far, so good. However, the devil is in the details. On careful inspection, it turns out that the synapomorphies of Node 4 are entirely composed of post-cranial characters. There is absolutely no anatomical support whatsoever for placing Paliguana, an isolated skull, at Node 4. Node 3 makes good sense. It consists most significantly of characters relating to the quadrate which, in turn, govern the ability of the jaw to exert pressure when closed. This is a very significant evolutionary development which is discussed at length by Carroll (1988) and in the main entry under Lepidosauriformes. However, Node 2 has just 1 synapomorphy: the medial centrale is significantly larger than the the lateral centrale -- a character of the "hand" which seems very easily influenced by local environmental conditions. Paleagama could not be scored on this character, nor could it be scored on any of the node 3 synapomorphies. Thus, there is also no support for placing Paleagama where it is, either. In short, just as Carroll (1988a) argued, there is no basis for separating Paliguana, Saurosternon, and perhaps even Paleagama. Combined, they give an interesting and useful description of a grade or hypothetical ancestral proto-lepidosaur, which one might call "Paliguanidae" so long as the monophyly of the group were taken with a grain of salt.
So why have the Paliguanidae been banished from the Notes? For several reasons. First, while there is no basis for separating these taxa, there is also no basis for uniting them in a phylogenetic context. More importantly, there is no reason to treat them as distinct phylogenetic entities at all, together or separately. As Feduccia 1999) has pointed out, there is not a great deal of utility in giving every scrap of lithified bone a Latin binomial. Unless there is enough information to place the fossil in a reasonably secure phylogenetic context, it makes no sense for phylogenetic taxonomy to recognize the fossil taxonomically. This simply leads to absurd results -- such as placing an isolated head in a category described only by post-cranial characters -- and cluttering cladograms with meaningless branches.
This is not to say that the paliguanids should be sold as paperweights. They are basal lepidosauriforms, or in that general neighborhood. They already have a taxonomic name: "Lepidosauriformes incertae sedis." If we are willing, as good cladists, to give up the idea of looking to nodes for an actual ancestor, then there is no reason not to refer a specimen to a node or stem group. The practice of refusing to do so reflects (a) a reaction to the Linnaean taxonomy's insistence on ancestry and (b) the sound logical principle that we can never be certain that a specimen is actually the last common ancestor of any two other specimens. However, a node is not only the LCA, but all of its descendants. All we know about the LCA are its synapomorphies. If a specimen has these characteristics, and has no characters which would render it unique, it should be referred to the node, and to no other included taxon, pending a more subtle analysis or additional material.
The same analysis applies to stem groups, such as Lepidosauromorpha = all Sauria closer to lizards than to birds, and Lepidosauriformes = all Lepidosauromorphs closer to lizards than to plesioraurs. Paliguana and Saurosternon are clearly that. But, as we have seen, they are no more than that. Thus, they are appropriately referred to the stem group, incertae sedis if one must; but, perhaps better, simply Lepidosauriformes. This is not an argument against rigor. Paleagama apparently lacks the synapomorphies for nodes 1, 2 or 3. At best, it is a diapsid.
In the last analysis Gauthier et al. (1988) and Carroll (1988a) are both correct. The exclusion of Paleagama has little impact on Carroll's (1988 & 1988a) hypothetical proto-Lepidosaur, and this construct forms a useful and appropriate model. However, there is no "Paliguanidae." The head and body can be analyzed together -- as a description of the basal Lepidosauriform morphotype.
Lepidosauriformes: Paliguana, Saurosternon. Lizards > plesiosaurs.
from the Late Permian?
Phylogeny: Lepidosauromorpha: Sauropterygia + *. Kuehneosauridae + (Marmoretta + (Sophineta + Lepidosauria)).
Characters: $ quadrate wide and bowed in lateral view; $ retroarticular process large. Stem species intermediate between basic diapsid form and lepidosaurs: lower temporal bar absent; frontal-parietal suture apparantly fairly loose on skull roof; quadrate articulation with pterygoid flexible or absent; dorsal end of quadrate enlarged to articulate with squamosal (streptostyly); no fenestration of scapulocoracoid or pelvis; vertebral centra amphicoelous; space for jaw musculature limited, but some kinesis acheived and development of space for pterygoideus allows force to be exerted by jaws when mouth is closed.
References: Carroll 1988a); Clark & Hernandez 1994); Gauthier et al. 1988).
Image: sketch after Carroll 1988). For purposes of the essay, note that this is all of Paliguana. ATW000301.
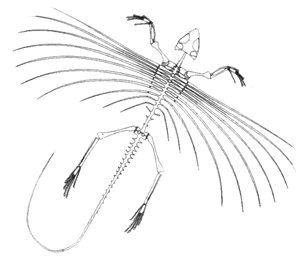 Kuehneosauridae: Kuehneosaurus, Icarosaurus.
Kuehneosauridae: Kuehneosaurus, Icarosaurus.
Late Triassic of North America & Europe
Lepidosauriformes: (Marmoretta + (Sophineta + Lepidosauria) + *.
Unclear whether this is the sister of Lepidosauria or Sphenodontidae. Possibly a garbage taxon of basal Lepidosauria with some but not all lizard synapomorphies. Carroll includes Paliguanidae in this group.
Image: Icarosaurus from La scoperta dell'Icarosaurus.
Links: reptiles; Nathis Fauna Reptilelen Slangen en hagedissen (Dutch); Kuehneosaurus; Paleontology and Geology Glossary: K; Dinosaurios: Kuehneosaurus; A NEW TRIASSIC GLIDING REPTILE FROM THE DANVILLE BASIN; Chapter 14 The Evolution of Flight; La scoperta dell'Icarosaurus (Italian); Era Mesozoica Triassico - I Paracadutisti (Italian). ATW000226.
checked ATW031006, last revised MAK100929