Neusticosaurus edwardsii. Middle Triassic - Tethys Sea; length about 60 cm.
| Sauropterygia | ||
| The Vertebrates | Pachypleurosauridae |
| Vertebrates Home | Vertebrate | Vertebrate |
|
Abbreviated Dendrogram
Diapsida
├─Archosauromorpha
│
└─Lepidosauromorpha
├─Sauropterygia
│ ├─Placodontia
│ ├─Pachypleurosauridae
│ │ ├─Hanosaurus
│ │ └─┬─┬─Dactylosaurus
│ │ │ └─Anarosaurus
│ │ └─Serpianosaurus
│ │ └─Neusticosaurus
│ └─Eusauropterygia
│ ├─Nothosauridae
│ └─Plesiosauria
│ ├─Pliosauroidea
│ └─Plesiosauroidea
│ ├─Elasmosauridae
│ └─Cryptocleidoidea
└─Lepidosauriformes
├─Sphenodontia
└─Squamata
|
Contents
Overview |
|
|
Neusticosaurus edwardsii. Middle Triassic - Tethys Sea; length about 60 cm. |
The pachypleurosaurs were small, lightly built lizard-like reptiles, generally less than a meter (the smallest about 20 cm). They were clearly related to the nothosaurs, with which they were contemporary. For a long time the pachypleurosaurs were included among the nothosaurs, and originally even placed in the same family Nothosauridae. It is now coming to be realized that they represent a distinct group that was only related via a primitive sauropterygian common ancestor.
The neck is long but, unlike the Nothosauridae, the head is very small (like Claudiosaurus), so pachypleurosaurs probably fed on very small fish and Crustacea. Relatively small dorsal temporal openings in the skull also clearly distinguish the pachypleurosaurs from the nothosaur genera (and their close relatives) such as Lariosaurus, Ceresiosaurus, Corosaurus, Paranothosaurus and Simosaurus, in all of which the upper temporal openings are much larger than the orbits. The tail is deep and would have been used as a swimming organ. The legs were semi-flattened to serve as paddles, but the hips and shoulders were still powerful and functional enough to provide support on land. It is likely that these animals were able to crawl about on land like modern seals, dragging themselves by their strong forelimbs. As with the nothosaurids they were semi-aquatic forms, mostly lagoonal but also venturing out to sea, perhaps not unlike the modern marine iguana in that respect. Their small size meant however they probably did not venture out far, being primarily near-shore forms. MAK991211.
in all of which the upper temporal openings are much larger than the orbits. The tail is deep and would have been used as a swimming organ. The legs were semi-flattened to serve as paddles, but the hips and shoulders were still powerful and functional enough to provide support on land. It is likely that these animals were able to crawl about on land like modern seals, dragging themselves by their strong forelimbs. As with the nothosaurids they were semi-aquatic forms, mostly lagoonal but also venturing out to sea, perhaps not unlike the modern marine iguana in that respect. Their small size meant however they probably did not venture out far, being primarily near-shore forms. MAK991211.
Pachypleurosaurs are often portrayed as a monophyletic clade (Rieppel, 1989, Lui et al 2011) and sister group to the Eusauropterygia (Storrs 1993, Rieppel, 2000). The problem with this approach, as with Guathier's groundbreaking work on the theropod dinosaurs (Gauthier 1986), is that they don't take into account reversals (loss of shared character traits (synapomorhies)). convergences can also provide a misleading signal. Such cladograms produce hypotheses where there is an infinite regress of sister taxa and ghost lineages, in which no ancestor-descendent relationship can ever be recovered. Statistical parsimony analsyes give variying ersults, but often show these monophyletic ancestrors to in fact be a paraphyletic grouping of early ancestral types and persistantly primitive later types, as happened with the Ceratosauria; and with the pachypleurosauria where for example Keichousaurus is now understood as basal nothosaur Holmes et al 2008. Similarily, Wu et al. (2011) restudy, using new material, of the abarrent basal pachypleurosaur Wumengosaurus reject Jiang et al (2008) (who originally named the species) original assement of a monophyletic Pachypleurosauria, replacing it with a clade that includes the Nothosauroidea and those taxa assigned to the paraphyletic Pachypleurosauria.
The nothosaurs themselves still tend to be understood monophyletically, although they raise the same problem of ancestor-descendant relationship as they are intermediate in organisation between pachypleurosaurs and higher sauropterygia (pistosaurs and plesiosaurs). Here it is worth considering the possibility of a basal nothosaur like Simosaurus as ancestral to both specialised nothosaurs and plesiosaurs. Alternatively the pistosaurs and plesiosaurs may be derived from a primitive pachypleurosaur-like ancestor such as Corosaurus. MAK111125
Finally, long bone histological studies of Triassic sauropterygia reveal two quite different bone growth patterns, and imply a paraphyletic or even polyphyletic pachypleurosauria, quite distinct from the conventional gross-morphology based tree topologies (Klein 2010). Although one may question how much undistorted phylogenetic signal such studies provide, since various functional factors come into play as well (Cubo et al 2005), this is certainly one more factor to key into the overall picture.
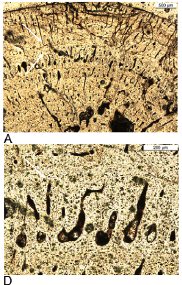
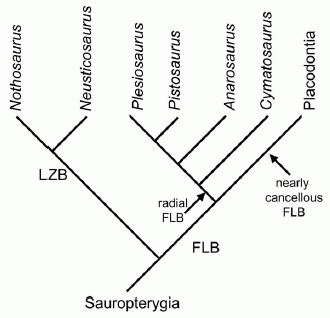 Phylogeny based on bone tissue types in Sauropterygia long bones, which differ markedly from standard tree topologies. Taking only bone histological data, pachypleurosaurs are polyphyletic because Neusticosaurus shows lamellar-zonal bone whereas Anarosaurus heterodontus has incipient fibrolamellar bone. Cymatosaurus already shows a radial vascular pattern and distinct layers of fibrolamellar bone within the parallel-fibered bone tissue. Placodonts have the most advanced bone tissue within Sauropterygia. LZB = Lamellar-zonal bone tissue type, FLB = Fibrolamellar bone tissue type. This hypothesis is not yet supported by a data matrix or phylogenetic analysis and thus remains provisional. Diagram and caption from Klein 2010, Creative Commons attribution license. |
We think of the pachypleurosaurs as pretty uniform. All share a common long-necked and small headed but otherwise standard lizard-like form. The long neck itself may be a common aquatic adaptation, as it is also found in prolacertiformes and choristoderans. It seems however that these creatures were very metabolically diverse, as three distinct bone histologies are known (Klein 2010). This diversity dates back even to the early-middle Triassic (Anisian age) indicating a rapid evolutionary radiation. The two main types are:
The lamellar-zonal bone (LZB) type, the standard low metabolic reptilian type, which is the primtive or plesiomorphic condition for amniotes. It consists of lamellar bone with only low to moderate vascularization, and bone tissue is dominated by longitudinally arranged simple vascular canals (Klein 2010 p.3). Growth tends to be sporadic, indicating ectothermic metabolism. This bone type is associated with lepidosaurs, crocodiles, and turtles This is found in the Serpianosaurus-Neusticosaurus lineage (Hugi et al 2011). Surprisingly, nothosaurs (Nothosaurus (Klein 2010) and Ceresiosaurus (Hugi 2011) also shows this typical lamellar-zonal bone type, but not more advanced forms such as Pistosaurs. The implication may be that these two are related, and that nothosaurids evolved distinct from other higher sauropterygia ( (Klein 2010).
The fibrolamellar bone (FLB) type in contrast involves a complex bone structure deposited very rapidly. It is highly vascularized (many blood vessels) and dominated by radial, laminar or irregularly arranged vascular canals. The FLB type is produced by fast growth rates. It always occurs in association high metabolic rates, and is typical of dinosaurs, extant birds, and large mammals. Growth is not only rapid but also constant, as evident in sauropods, large herbivorous mammals, and most birds. However, growth can also be cyclically interrupted, as in the case of many nonavian dinosaurs. (Klein 2010 p.3, see further references therein). Among sauropterygia, it is found in the pachypleurosaur Anarosaurus and in pistosaurs such as the basal Cymatosaurus and the more derived Pistosaurus, as well as in the crown group Plesiosauria (plesiosauroids and pliosauroids). What is extraordinary is that morphologically Anarosaurus is a very basal pachypleurosaur; we see here a disjunction between gross morphology and metabolism, which reminds us of the need for a total evidence approach.
The fibrolamellar pachypleurosaurs themselves are divided into two further groups, referred to in Klein 2010 as histotype A and histotype B. Both are dominated by a radial pattern of vascularization. However, vascular density is distinctly higher in histotype B. On average, vascular canals take up between 5% to 10% of histotype A bone tissue, but between 10% to nearly 20% in histotype B. Histotype B is associated with Anarosaurus, whereas, curiously, the somewhat less metabolically intense histotype A (although still superior to the nothosaur condition) is assocuiated with the ore advanced form, Cymatosaurus, although this association si tentative because Cymatosaurus is not known with certainty from postccranial material. In any case, this indictaes a whole range of different metabolic types evolving alongside each other. Finally, a sample of placodont bone vascular density is even higher than in histotype B, dominated by very large longitudinal canals, indicating an even higher level of metabolism. (Klein 2010)
|
If fibrolamellar bone was inherited by the Sauropterygia from a terrestrial ancestor, this could place their origin in the Archosauria, where fibrolamellar growth appears to be basic (Ricqles et al 2008 cited in Klein 2010) rather than the Lepidosauromorpha. Ironically, this would then enable Chelonii (turtles) to be both related to Sauropterygia (as proposed by Rieppel and others) and nested in the archosaurs (as molecular phylogeny seems to show). The problem is that this goes as much against all available morphology as crown archosaurian turtles do; if sauropterygians are archosauromorphs they much more likely branched off at a very basal level [1] On the other hand, the fact that Neusticosaurus and Nothosaurus show the more primitive, lamellar-zonal bone structure, supports the hypothesis of Lepidosauromorph origin (Lepidosauromorpha having retained the basal amniote lamellar-zonal condition) and implying that fibrolamellar bone developed multiple times within the Sauropterygia (Klein 2010 p.23).
It is however very unlikely that small lizard-like pachypleurosaurs were endotherms or in any way metaboloically similar to dinosaurs, birds, or mammals. There are limits to the reliability of such studies as metabolic indicators; Haversian or fibrolamellar bone is fairly common in young crocodilians and sometimes found in adults, it has also been found in turtles and tortoises, and temnospondyl amphibians, but is often absent in small birds, bats, shrews and rodents. (Enlow 1963, Reid 1984, Reid 1997 cited via Wikipedia, Ray et al 2009). In all other respects, pachypleurosaurs were classic reptiles with elongated bodies and broadly lizard-like appearance, lacking for example the long limbs and parasaggital (erect) gait of mammals and higher archosaurs (rauisuchia, dinosaurs, and birds). Endothermy would also have been a distinct disadvantage for a small aquatic animal due to loss of body heat. However, fibrolamellar bone does indicate rapid growth (Ray et al 2009). We can assume then that the sauropterygia adopted a number of distinct growth strategies. At one extreme there was the slow growing and classic reptilian regime of the neusticosaurs and nothosaurs; on the other the ultra-rapid growth of the anarosaurs and placodonts, which had an even higher metabolic growth rate when young than the pistosaurs and plesiosaurs. Once adult size was reached, anarosaurs and placodonts probably settled down to a typically reptilian liefstyle.
The question is, how much did these ontogenic, metabolic, and physiological processes factor in to phylogeny? On the basis of histological data alone, most current sauropterygian clades - Nothosauria, Pachypleurosauridae and Eusauropterygia - would be rejected, although the higher Pistosauroidea and the Plesiosauria are retained. Neusticosaurus is now related to Nothosaurus, whereas the basal pachypleurosaur Anarosaurus is relocated to the pistosaur clade, all of which gives a very different dendrogram topology. Further studies are required, ideally incorporating gross morphological, histological, and stratographic or stratocladistic methodologies, in order to arive at a better understanding of early sauropterygian phylogeny. MAK111101
Notes:
[1]John Merck favours thalattosaurs and Euryapsida (Ichthyopterygia + Sauropterygia) as the sister group to all other archosauromorphs (referenced in DML posting (Tom Holtz), see also Tom Holtz webpage) Cladograms given by Borsuk-Białynicka & Evans, 2009 using using the data matrix of Müller 2004 also present thalattosaurs, ichthyosaurs and sauropterygia as basal archosauromorphs. Because of the uncertainty regarding the placement of these groups we have retained more traditional phylogenies, at least until further research confuiirms their archosauromorphian status
Pachypleurosauridae: Nopsca 1928. Neusticosaurus Seeley, 1882; Anarosaurus Dames, 1882; Dactylosaurus Bardet, 1992; Hanosaurus Lee, 1958; Serpianosaurus Rieppel 1989. Pachypleurosaurus and Phrygosaurus are considered nomina dubia, and have been sunk into Neusticosaurus and Serepianosaurus, respectively.
Range: Middle Triassic (possibly Early Triassic in China) (possibly to Late Triassic [CG85] [RK95] in Europe) of Central Europe & China. [R95].
Nothosauria :Eusauropterygia+ * : Hanosaurus + ((Dactylosaurus + Anarosaurus) + Serpianosaurus + Neusticosaurus)).
 Characters: S-M size (<150 cm) [CG85]; similar to nothosaurs; skull unusually small [CG85]; skull not depressed as in Nothosauridae [R89]; prefrontal broad & domed [RK95]; frontals with concave lateral margin participating in orbit [RK95]; postorbital skull shorter than preorbital [R89] [RK95]; upper fenestra smaller than large orbit [CG85]; strong upper bar; no lower temporal bar [CG85]; UTF usually small [R89]; postorbital primitively enters margin of UTF [R89]; quadrate embayed (impedence-matching ear for airborne vibrations & thus amphibious?) primitive) [R89] [RK95]; jaw joint not behind occipital condyle [R89] [RK95]; pterygoids expanded to completely cover braincase ventrally [CG85]; ectopterygoid absent [R89] [RK95] (contra [CG85]); jaws not elongated; jaw with retroarticular process bearing dorsal trough [RK95] [3]; anterior teeth nor strongly procumbent [RK95]; vertebrae amphicoelous throughout [R89]; long 18 cervicals) neck; dorsal vertebrae have low neural spines, thickened and dense (pachyostosis
-- see Note); pachyostosis extends to ribs in more derived species; 3 sacral vertebrae; dorsoventral extension of vertebral spines near base of tail; girdles poorly ossified (thus obligate marine?); limbs not highly derived for aquatic life; sexual dimorphism in forelimb [ORS99]; interclavicle, posterior process rudimentary or absent [RK95]; radius slightly longer than ulna [RK95]; juveniles with entepicondylar foramen distal edge of ossified portion of humerus [R95]; iliac blade reduced to narrow dorsal process [RK95]; restricted to epicontinental & coastal areas. Appear to have originated in China and migrated to Europe via the northern border of Tethys Sea [RK95]. ATW070113.
Characters: S-M size (<150 cm) [CG85]; similar to nothosaurs; skull unusually small [CG85]; skull not depressed as in Nothosauridae [R89]; prefrontal broad & domed [RK95]; frontals with concave lateral margin participating in orbit [RK95]; postorbital skull shorter than preorbital [R89] [RK95]; upper fenestra smaller than large orbit [CG85]; strong upper bar; no lower temporal bar [CG85]; UTF usually small [R89]; postorbital primitively enters margin of UTF [R89]; quadrate embayed (impedence-matching ear for airborne vibrations & thus amphibious?) primitive) [R89] [RK95]; jaw joint not behind occipital condyle [R89] [RK95]; pterygoids expanded to completely cover braincase ventrally [CG85]; ectopterygoid absent [R89] [RK95] (contra [CG85]); jaws not elongated; jaw with retroarticular process bearing dorsal trough [RK95] [3]; anterior teeth nor strongly procumbent [RK95]; vertebrae amphicoelous throughout [R89]; long 18 cervicals) neck; dorsal vertebrae have low neural spines, thickened and dense (pachyostosis
-- see Note); pachyostosis extends to ribs in more derived species; 3 sacral vertebrae; dorsoventral extension of vertebral spines near base of tail; girdles poorly ossified (thus obligate marine?); limbs not highly derived for aquatic life; sexual dimorphism in forelimb [ORS99]; interclavicle, posterior process rudimentary or absent [RK95]; radius slightly longer than ulna [RK95]; juveniles with entepicondylar foramen distal edge of ossified portion of humerus [R95]; iliac blade reduced to narrow dorsal process [RK95]; restricted to epicontinental & coastal areas. Appear to have originated in China and migrated to Europe via the northern border of Tethys Sea [RK95]. ATW070113.
Comments: Pachypleurosaurs are small (generally less than a meter), primitive, superficially lizard-like forms. Near-shore, coastal dwellers, they preferred equatorial regions and flourished briefly from the early Middle Triassic (Early Anisian) to the early Late Triassic (early Carnian). In size and appearance they were not very different to earlier Permo-Triassic diapsids like Claudiosaurus, from which they may have evolved (see note on conflicting sauropterygia hypotheses). Although specialised in a number of features, which has led to them being considered a monophyletic clade, they are the obvious ancestors for all other sauropterygians, and indeed, the only possible ancestral forms in the fossil record from which the other types could have evolved (through increase in size and loss of specialised features). Metabolically diverse, bone histology shows that they included both slow growing and fast growing lineages. MAK111125
Notes: [1] Pachyostosis, the thickening of bones, is common in recently aquatic tetrapods. Examples include mammalian Sirenia e.g. manatees) and an unusual plesiosaur, Pachycostasaurus. deRicqlès 1989). But see Gaudant & Meunier (1996). This may have more to do with ballast for a benthic habit.
[2] The current belief is that the pachypleurosaur line forms part of the bridge between the Sauropterygian stem (e.g. placodonts) and the Eusauropterygia e.g. plesiosaurs). The connection is an admittedly difficult one to make, but the fossil record gives few options. To make this phylogenetic region yet stranger, a prolonged stare at the pachypleurosaurs tends to bring turtles to
 mind -- yet another taxon of still-uncertain affinities. Look, for example, at the way the dorsal and caudal ribs seem to rise up above the level of the sacrum (the fused area of bone at the intersection of the pelvis and the spine) in some photographs of the nothosaur, Keichousaurus. Turtles are the only group of vertebrates with both sets of limb girdles inside the rib cage. Yet it would take little modification to give this unique character to a pachypleurosaur. The pachyostosis of the ribs, from which the group derived its name, is similar to the broad, heavy ribs of turtles. Although pachypleurosaurs themselves are unarmored, one group of placodonts has strikingly turtle-like armor. This relationship between sauropterygians and turtles was widely defended at the end of the last century. It seemed to fall apart, on details of skull anatomy among other things. However, the concept has recently been revived by Olivier Rieppel and others. The idea has also gained some support from DNA cladistics which tends to place the turtles up above the diapsid split, although their placement is sometimes (perhaps embarrassingly) closer to archosaurs than to lepidosaurs. However, given the total absence of (other?) living sauropterygians, a little uncertainty is only to be expected.
mind -- yet another taxon of still-uncertain affinities. Look, for example, at the way the dorsal and caudal ribs seem to rise up above the level of the sacrum (the fused area of bone at the intersection of the pelvis and the spine) in some photographs of the nothosaur, Keichousaurus. Turtles are the only group of vertebrates with both sets of limb girdles inside the rib cage. Yet it would take little modification to give this unique character to a pachypleurosaur. The pachyostosis of the ribs, from which the group derived its name, is similar to the broad, heavy ribs of turtles. Although pachypleurosaurs themselves are unarmored, one group of placodonts has strikingly turtle-like armor. This relationship between sauropterygians and turtles was widely defended at the end of the last century. It seemed to fall apart, on details of skull anatomy among other things. However, the concept has recently been revived by Olivier Rieppel and others. The idea has also gained some support from DNA cladistics which tends to place the turtles up above the diapsid split, although their placement is sometimes (perhaps embarrassingly) closer to archosaurs than to lepidosaurs. However, given the total absence of (other?) living sauropterygians, a little uncertainty is only to be expected.
[3] The "trough" may be the same as what Sander (1989) identifies as a pit to receive the posterior process of the quadrate in Neusticosaurus. See Some Jaw Notes.
References: Carroll & Gaskill (1985) [CG85]; O'Keefe et al. 1999) [ORS99]; Rieppel (1989) [R89]; Rieppel (1995) [R95]; Rieppel & Hagdorn (1997); Rieppel & Kebang (1995) [RK95]. ATW070113.
Further note A monophyletic Pachypleurosauridae is supported by Rieppel 1989 Storrs 1993, Rieppel 1998 and Jiang et al 2008 but rejected by Holmes et al 2008 and Wu et al. 2011. Perhaps a distinction could be made between what seem to be a paraphyletic assemblage or evolutionary grade of basal forms and specialised monophyletic lineages of more typical pachypleurosaurs MAK111125
Hanosaurus: H. hupehensis Young 1972
Range: Middle Triassic (Anisian) Jialingjiang Formation of Nanzhang, Hubei province, China
Phylogeny: Pachypleurosauridae : (Dactylosaurus + Anarosaurus) + (Serpianosaurus + Neusticosaurus)) + *.
Characters: elongate skull with long posterior orbits, no lower temporal opening, elongate supratemporal opening (Lucas, 2001,p p.115-6)
Comments: known from an incomplete skull and some postcrania, and originally interpreted as a thalattosaur, cladistic analysis by Reippel 1998 shows this species to be a basal pachypleurosaur close to Keichousaurus, and supports a sister group relationship between thalattosaurs and sauropterygia. This was further supported by Li et al 2011 who also make the Saurosphargidae the sister group of the Thalattosauriformes. However Holmes et al 2008's analysis presents Keichousaurus as a very primitive (basal in cladistic speak) nothosaur, and hence the the pachypleurosaurs as paraphyletic. It may still be that Keichousaurus and Hanosaurus are closely related, with one evolving to the nothosaurian condition, another to the pachypleurosaur state, although such ancestor-descendent interpretations fall outside cladistic methodology, which is based on phylogenetic hypotheses limited solely to sister group relationships MAK111125
checked ATW090306, revised MAK111125
Using this material. All material by ATW is public domain and may be freely used in any way (also any material jointly written by ATW and MAK). All material by MAK is licensed Creative Commons Attribution License Version 3.0, and may be freely used provided acknowedgement is given. All Wikipedia material is either Gnu Open Source or Creative Commons (see original Wikipedia page for details). Other graphics are copyright their respective owners