 Taxa on This Page
Taxa on This Page| Sauropterygia | ||
| The Vertebrates | Sauropterygia |
| Vertebrates Home | Vertebrate | Vertebrate |
|
Abbreviated Dendrogram
Diapsida
├─Archosauromorpha
│
└─Lepidosauromorpha
├─Sauropterygia
│ ├─Placodontia
│ └─Nothosauria
│ ├─Wumengosaurus
│ ╞═Pachypleurosauridae
│ └─┬─Nothosauridae
│ └─Plesiosauria
│ ├─Pliosauroidea
│ └─Plesiosauroidea
│ ├─Elasmosauridae
│ └─Cryptocleidoidea
└─Lepidosauriformes
├─Sphenodontia
└─Squamata
|
Contents
Overview |
Taxa on This PageThe Sauropterygia or "lizard flippers" are an important group which includes a wide range of aquatic lizard-like, crocodile-like, long-necked "Loch Ness monster"-like, and large-headed killer whale-like aquatic reptiles. Among the great prehistoric success stories, they first appeared at (or just before) the very start of the Mesozoic and flourished right until the end of the era. Yet as with the Ichthyosauria, they are so specialized their ancestry and evolutionary relationships are still somewhat obscure (see comments on Sauropterygian origins.
Earlier forms (pachypleurosaurs) were mostly small amphibious lizard-like animals with slender, elongated limbs and small heads, that lived in estuarine and near-shore environments. They quickly increased to several meters in size and ventured further out to sea, although still retaining amphibious traits and sinuous lizard-like form (nothosaurs). By the Middle Triassic there were a number of divergent lineages. In addition to the line leading to the successful Jurassic plesiosaurs, there were several divergent branches of early sauropterygians, including pachypleurosaurs, several lines of nothosaurs, and the transitional pistosaurs. These early divergent branches show successive stages in adaptation to oceanic life.
The Carnian extinction took out most of these early Sauropterygia, but the one surviving group, the plesiosaurs, underwent a tremendous evolutionary radiation around the very end of the Triassic period and the very start of the Jurassic, diverging rapidly into variously lineages in a typical example of a macroevolutionary adaptive radiation. It is not yet clear whether this event coincided with and was caused by the full development of the plesiosaur locomotor plan, or whether it was simply opportunistic factors, such as the mass extinction of potential competitors in the Rhaetian turnover, that allowed the plesiosaurs to exploit the potential of a previously evolved locomotor plan (see below). In any case by the early Jurassic there were a diverse range of long- medium- and short-necked forms, some with small heads others with large. These were all open ocean types, completely adapted for marine life, and usually 3 metres or more in length. The two principal morphs were the plesiosaurs (long necked "Loch Ness monster" types) and pliosaurs (reptilian killer whales). However, the evolutionary relationships of the Plesiosauria are complex, and it appears that long-necked and short-necked types occurred in a number of lineages.

The Sauropterygia are united by unique derived features features of the skull and shoulder girdle. As Carroll & Gaskill (1985) note, the fundamental relations of the shoulder girdle elements are altered in the Sauropterygia. The figure illustrates the differences in shoulder structure between a basal amniote (adapted from Carroll, 1988) and the basal sauropterygian, Neusticosaurus (adapted from Carroll & Gaskill 1985). In the terrestrial amniote, force is directed by the humerus against the plate-like scapulocoracoid, which dissipates the force over a wide area and is, in turn, supported by extensive contact with the dermal bones of the clavicle and interclavicle. In the sauropterygian shoulder, a direct, medially directed force would probably cause the shoulder girdle to collapse. However, the anterior and posterior strains caused by paddling and steering simply bind the girdle more strongly together by forcing the interclavicles and coracoids against their opposite numbers. The fundamental re-engineering of the shoulder girdles suggests that the strains involved were considerable, i.e., that the sauropterygians were fast, agile swimmers.
A remarkably similar arrangement was evolved in the pelvic girdle of derived sauropterygians, e.g. plesiosaurs, and both girdles were connected by a net of gastralia (stomach ribs) probably reinforced by stout ligaments. This arrangement and the problems of plesiosaur locomotion, will be covered on a subsequent page. MAK991206, ATW061220.
Sauropterygia: definition variable in the literature. Used here as placodonts + plesiosaurs.
Range: Early Triassic to Late Cretaceous.
Postorbital skull elongated; upper fenestra enlarged; "euryapsid" (lower temporal arch lost) [CG85]; quadrate rigid (primitive) [CG85]; elongate neck (>7 cervicals); limbs relatively large [CG85]; curved humerus; radius equal length to ulna.
ATW061216.
Comments: The Sauropterygian can be divided into six main types. The Pachypleurosauridae represent an ancestral lineage or series of lineages, all are morphologically quite similar, small lizard-like near shore forms, they include both slow and fast growing types. The Placodontia are highly specialised armoured turtle like forms with variable diet (molluscivore, seaweed, filter feeding, etc). The Nothosauridae are larger forms otherwise resembling the pachypleurosaurs. The Pistosauridae are intermediate between pachypleurposaurs and nothosaurs on the one hand, and crown group sauropterufgia (Plesiosauria) on the other. The Pliosauroidea started out as small to medium sized long-necked and small headed types, but later became medium to very large short-necked and large headed forms. These were the superpreditors of the middle Jurassic through to mid Cretaceous, immortalised in the popular geek consciousness as the mythical 25 meter Liopleurodon of Walking With Dinosaurs fame (in fact 8 to 12 meters was more realistc, still a very respectable size for an apex predator). Finally the Plesiosauroidea are your standard "loch ness monster" or "antediluvian sea monster", with a small head on a long sinuous neck, and a broad turtle like body. Some advanced forms however became secondarily pliosaur-like. MAK111125
Sauropterygian origins: At one time included with the Ichthyosuria, polyphyletic Protorosauria (Protorosaurus + Araeoscelida), and a seperate Placodontia in the Subclass Euryapsida (Colbert, 1969); the Placodonts were then given their own subclass Kuhn 1969. Later Carroll 1981 derived the pachypleurosaurs, and hence the Sauropterygia as a whole, from the late Permian diapsid Claudiosaurus. Claudiosaurus itself he placed in the Lepidosauromorpha. Other workers however considered Claudiosaurus a more primitive form, predating the Archosaur-Lepidosaur divide (e.g. Benton 1985, see Diapsid phylogeny) , and this is the current consensus position which we have followed here. Rieppel on cladistic grounds returned to Sauropterygia to the Lepidosauromorpha (minus Claudiosaurus), and made them the sister group of the Placodontia, thus revising in part the Euryapsida; alternatively the Placodontia are included in the Sauropterygia, but distinct from pachypleurosaurs + (nothosaurs + (plesiosaurs))). More recently John Merck returned the Ichthyosaurs to the Euryapsida as (Thalattosauria + Euryapsida (Ichthyopterygia + Sauropterygia), but moved them from basal lepidosauromorpha to basal archosauromorphs (ref: DML posting (Tom Holtz). This gives three possible, and perhaps equally plausible, hypotheses for sauropterygian origins: as stem neodiapsids derived from Claudiosaurus or a similar form, as stem lepidosauromorphs, or as stem archosauromorphs. MAK111102
Links: Sauropterygia -- The Dinosauricon; Sauropterygia; Turtle Origins; Literature - Sauropterygia; muschelkalk lipperland (German: never saw this site before. Very nice.); ???????? Japanese); Untitled Document; Literature - Sauropterygia; Sauropterygia after Rieppel, 1997, 1998ab and Rieppel & Hagdorn, 1997; Fossil Room; some additions to Re: MOSASAURS AND PLIOSAURS; MOSASAURS AND PLIOSAURS; muschelkalk lipperland (German).
References: Carroll & Gaskill 1985) [CG85]. ATW061216.
| Changing classifications of the Sauropterygia | ||
| Colbert, 1969: | Carroll 1988: | Benton 2005: |
| Subclass Euryapsida Order Protorosauria Order Ichthyosauria Order Sauropterygia Suborder Nothosauria Suborder Plesiosauria Superfamily Pistosauroidea Superfamily Pliosauroidea Superfamily Plesiosauroidea Order Placodontia Subclass Diapsida |
Subclass Diapsida Order Placodontia (incertae sedis) Infraclass Lepidosauromorpha Superorder Lepidosauria Superorder Sauropterygia nbsp; Family Claudiosauridae (incertae sedis) Order Nothosauria Order Plesiosauria Superfamily Pliosauroidea Superfamily Plesiosauroidea Infraclass Archosauromorpha |
Subclass Diapsida Order Araeoscelidia Infraclass Ichthyosauria sedis mutabilis Infraclass Lepidosauromorpha Superorder Lepidosauria Superorder Sauropterygia Order Placodontia Order Nothosauroidea Suborder Pachypleurosauria Suborder Nothosauria Order Plesiosauria Suborder Pliosauroidea Suborder Plesiosauroidea Infraclass Archosauromorpha |
The above represent Linnaean/Evolutionary rank-based classifications, which allow ancestral taxa, and should not be confused with Cladistic clade-based phylogenies which do not. Both methodologies are equally valid and complementary; it is not the case that one "disproves" the other. MAK111103
 Nothosauria: plesiosaurs > placodonts. This was traditionally the name for a paraphyletic group including pachypleurosaurs and nothosaurs. Since a lot of good work was done on the plesiomorphic state of the group, we insert it here as a stem group supporting Sauropterygia. Most references prior to about 1990 use "nothosaurs" to refer to this group, rather than to Nothosauridae.
Nothosauria: plesiosaurs > placodonts. This was traditionally the name for a paraphyletic group including pachypleurosaurs and nothosaurs. Since a lot of good work was done on the plesiomorphic state of the group, we insert it here as a stem group supporting Sauropterygia. Most references prior to about 1990 use "nothosaurs" to refer to this group, rather than to Nothosauridae.
Range: Early Triassic to End Cretaceous
Phylogeny: Sauropterygia : Placodontia + * : Pachypleurosauridae + Eusauropterygia. ATW061220
Characters: "euryapsid" (lower temporal arch lost) [CG85]; quadrate rigid (primitive) [CG85]; quadrate embayed [R89]; pterygoids usually meet ventrally covering braincase, occluding interpterygoid vacuities [CG85]; pterygoid with parasagittal ridges medial to LTF (possibly pterygoideus muscle) [CG85]; tooth implantation typically thecodont [R89]; increase in neck length by increasing number of vertebrae (but number in trunk is primitively almost constant) [CG85]; scapula and coracoid separately ossified [CG85]; cleithrum & sternum absent [CG85]; clavicles & interclavicle form solid bar anteriorly & ventrally [CG85]; large opening between medial surfaces of scapulae & coracoids [CG85]; ventral margin of scapula broadly underlying posterior clavicle (opposite of usual tetrapod arrangement) [CG85]; interclavicle small & ventral to clavicles [CG85] [1]; coracoid posterior to surface of glenoid [CG85]; humerus remains relatively straight [R89]. ATW061220
Comment: the phylogenetic position of the placodonts is unclear, due to their highly specialised nature and lack of obvious characteristics that can be compared with run of the mill sauropterygia. It is quite likely that they diverged later from pachypleurosaurs, in which case Nothosauria characteristics given here are simply plesiomorphic sauropterygian traits which are lost by ancestral placodonts. Assuming this is so (and there don't presently seem to be any peer review papers pushing thsi line, so it's just the present author on his soap box) Nothosauria would become a junior synonym for Sauropterygia. If Pachypleurosauria turn out to be paraphyletic as well, that would mean that Sauropterygia = Nothosauria = Pachypleurosauridae. See also comments on Wumengosaurus, below. MAK111125
Notes: [1] See images at Sauropterygia and Nesticosaurus.
References: Carroll & Gaskill (1985) [CG85]; Rieppel (1989) [R89]. ATW061220.
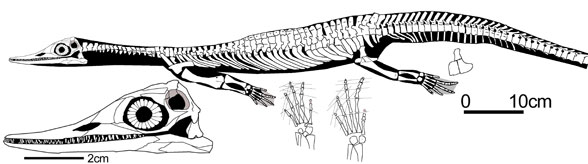
Wumengosaurus: W. delicatomandibularis Jiang et al 2008
Range: middle Anisian (Middle Triassic) of Panxian, Guizhou Province, southwestern China.
Phylogeny: Sauropterygia (= paraphyletic Pachypleurosauridae?) : Pachypleurosauridae + Placodontia + Nothosauridae + Pistosauroidea + *.
Characters: From the abstract of Jiang et al 2008: "unique specialization of the jaws that form an elongate pointed rostrum furnished with numerous small, monocuspid and vertically oriented teeth with a basally expanded crown and pointed tip, which is quite unlike any other sauropterygian known. Other diagnostic characters include: frontals paired; parietal unpaired; pineal foramen located centrally in broad parietal skull table; two carpal ossification; ilium with distinct preacetabular process at the base of the dorsally extending iliac blade; pubis plate-like and of rounded contours; two tarsal ossifications. The curved and distally expanded humerus, the reversed topological relationship of the clavicle and scapula, and the presence of three sacral ribs indicate sauropterygian affinities of the new taxon, whereas the 'butterfly-shaped' or 'cruciform' facet for the neural arch on the dorsal centrum surface indicate its eosauropterygian status."

Comments: The five known specimens show morphological variation, like that of Keichousaurus, Neusticosaurus, and Serpianosaurus. Originally considered by Jiang et al (2008) pachypleurosaurid with an unusually elongated rostrum, hence the species name. Wu et al. (2011) restudied this species is restudied on the basis of three new specimens. They revised Jiang et al (2008)'s analysis, rejected the idea of a monophyletic Pachypleurosauria, and questioned Wumengosaurus placement in the Pachypleurosauria. They suggest instead that Wumengosaurus might represent the basal-most member of a monophyletic clade that includes the Nothosauroidea and those taxa assigned to the paraphyletic Pachypleurosauria. David Peters tested Wumengosaurus against Stereosternum, Hupehsuchus, and Utatsusaurus and argued that they form a single clade with Wumengosaurus more derived than Stereosternum but more basal than Hupehsaurus, Utatsusaurus, and the Ichthyopterygia. The present author however feels that, whilst the taxa Peters lists are similar in appearance, this can be better explained in terms of convergence (Wumengosaurus being an ichthyosaur-mimicking sauropterygian rather than a proto-ichthyosaur. Similarily, if the basal placement of Placodonts and Wumengosaurus (rather like the original basal placement of Mesoauria in the Sauropsida prior to the Reptilia) is due to their specialised adaptations and loss of shared character traits (synapomorhies) rather than their never having those traits in the first place, the placodonts, Wumengosaurus, nothosaurs, and pistosaurs are all groups that evolved from an original pachypleurosaur ancestral assemblage (rather like the bubble-ancestor in romerograms), the details of which need to be still worked out. MAK111125
Graphic: skeletal reconstruction (right), David Peters, Reptile Evolution. Photo of fossil (left), MarineReptile.org (Triassic and Later Marine Vertebrate Symposium, Guanling, China)
Links: Wikipedia
revised ATW080328, checked ATW030715, revised MAK111125
Using this material. All material by ATW is public domain and may be freely used in any way (also any material jointly written by ATW and MAK). All material by MAK is licensed Creative Commons Attribution License Version 3.0, and may be freely used provided acknowedgement is given. All Wikipedia material is either Gnu Open Source or Creative Commons (see original Wikipedia page for details). Other graphics are copyright their respective owners