|
|
Sauropterygia |
| The Vertebrates |
Dactylosaurus
& Anarosaurus |
Sauropterygia: Pachypleurosauridae:
Dactylosaurus & Anarosaurus
Taxa on This Page
- Anarosaurus X
- Dactylosaurus X

The Cast System
Pachypleurosaurs did not fare well in World War II. A number of important museum collections in Europe were destroyed during this war, including holotypes in the collections of, for example, the Zoologisches Museum für Hamburg (now the Zoologisches Staatsinstitut und Zoologisches Museum), the Bristol City Museum, the Museo Civico di Storia Naturale di Milano, and Hiroshima University. In fact, from a historical perspective, the single most frequently-mentioned cause of destruction for vertebrate holotypes is bombing. And, of all wars fought in the last 200 years, World War II was the most destructive to fossils by a very large margin, due to the extensive, relatively unselective, bombing of metropolitan centers. See, for example, Benton et al. (2000) (holototype of Thecodontosaurus antiquus), Smith et al. (2001) (Stromer collection of Bahariya vertebrates), Leonard et al. (2005) (holotype of Lithornis), Dubrovo & Sanders (2000) (undescribed Patriocetus sp.), Grande & Bemis (1998) (holotype and only specimen of Amiopsis); Bennett 1996) (one of two specimens of "Gnathosaurus"), Damiani 2001) (only specimen of Cyclotosaurus ebrachiensis).
Dactylosaurus and Anarosaurus were the earliest European pachypleurosaurs. They are known only from very rare material excavated in Poland [1] and Germany. At the beginning of the war, the holotype of Anarosaurus was deposited in Göttingen, while two referred specimens were kept in Dresden and Jena, respectively. All three were destroyed. Thus, we have only casts made in the 1920s and
1930s. Dactylosaurus apparently differed from Anarosaurus in having much greater resistance to Allied bombing. No Dactylosaurus material was lost in the war. However, both known specimens of Dactylosaurus were natural molds to begin with.
Thus, both genera are now represented only by impressions. We are fortunate that pachypleurosaurs seem have had pleasant dispositions and thus left good impressions; and that their custodians had been disposed to impress a number of casts which were impressed upon various institutions for disposition in relatively bomb-proof structures, so that they may still be impressed for dispositive confirmation of the original published impressions, if one is so disposed .... But enough of that. The point, if any, of that extended linguistic exercise is that casts, like English words (and pachypleurosaurs for that matter), tend to diversify, accumulating small variations which may confuse interpretation.
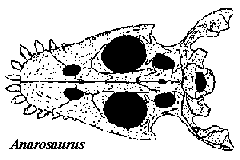 Anarosaurus pumilio was initially distinguished from other pachypleurosaurs based on some oddly spatulate anterior teeth and a suite of metric characters. However, Rieppel & Kebang [RK95] found that the various casts had warped a bit over the years, making the metric characters unverifiable. Worse, they were unable to verify the shape of the teeth. One cast showed some indications of this feature, but it also showed other variations from the type. They therefore referred it to a new species, A. heterodontus. This creates some difficulty, in that the new species is mainly distinguished by having a characteristic supposedly apomorphic for the old.
Anarosaurus pumilio was initially distinguished from other pachypleurosaurs based on some oddly spatulate anterior teeth and a suite of metric characters. However, Rieppel & Kebang [RK95] found that the various casts had warped a bit over the years, making the metric characters unverifiable. Worse, they were unable to verify the shape of the teeth. One cast showed some indications of this feature, but it also showed other variations from the type. They therefore referred it to a new species, A. heterodontus. This creates some difficulty, in that the new species is mainly distinguished by having a characteristic supposedly apomorphic for the old.
Dactylosaurus was initially believed to have two species: D. gracilis and D. schroederi, each represented by a single specimen. Rieppel & Kebang conclude that these are juvenile and adult of the same species, gracilis. That's well enough, but the holotype of the genus becomes the juvenile. This is seriously awkward, since the juvenile differs in material respects from the adult form. We are forced to admit that, as a consequence, we ultimately gave up on Dactylosaurus. ATW070125
[1] Dactylosaurus is from Upper Silesia -- part of Germany at the time it was discovered, but now Polish territory. The historical information related to both genera is taken from Rieppel & Kebang [RK95]
The European Pachypleurosaurs
In spite of our own failings, the best efforts of the RAF, the limited stratigraphic range of the fossils, and many other difficulties, the European pachypleurosaurs still make a really interesting little group of closely-related species. It is one of the few such systems for which one can expect meaningful answers to questions about evolutionary trends. To our knowledge, no one has turned over this particular rock to see what might lie under it. We lack the qualifications to do the job, but we couldn't resist the temptation to take a peek at a few of the sights.
The European pachypleurosaurs are Dactylosaurus, Anarosaurus, Serpianosaurus, and three species of Neusticosaurus: N. pusillus, N. peyeri, and N. edwardsii. They have little or no stratigraphic overlap and occur in the order listed. Rieppel & Kebang 1995); Sander (1989). So we're looking at a rather clean replacement series, with no reason to suspect extensive bushiness to the local phylogenetic tree. Undoubtedly, some bushiness existed. Mutations being random, divergence in all directions is to be expected. The wonder is that, over the roughly 10 million years during which pachypleurosaurs are found in Europe, many of the variations seem to be headed in a fairly uniform direction. That is, we seem to be looking at bona fide long-term trends based on natural selection.
We're not going to try and guess what factors were applying selective pressures. The point is that the critters don't seem to have specialized to subdivide their ecological niche. Rather they kept moving along a trajectory. It isn't easy to say what that trajectory accomplishes in a functional sense. Let's look at a few examples from the skull.

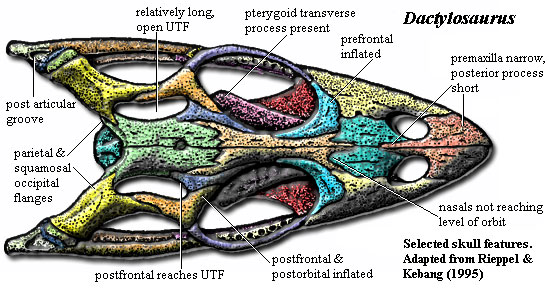
In general, the skull becomes more triangular. The posterior extension of the jaw becomes consistently shorter, and the jaw articulation is actually anterior to the occipital condyle in N. edwardsii. The entire postorbital region of the skull takes on an increasingly blocky or inflated appearance, with a corresponding increase in the relative size of the squamosal. The orbit generally becomes larger, and the jugal bar longer and less curved.
The posterior process of the premaxilla becomes longer and cuts more and more deeply between the nasals. Dactylosaurus has a small posterior process which separates the anterior ends of the nasals and extends no further than the posterior rim of the nares. Anarosaurus may even lack a posterior process, contrary to the trend. However, the four species of the Serpianosaurus-Neusticosaurus group show progressive elongation of the process so that, in N. edwardsii, the premaxilla reaches the frontals and may entirely eliminate the midline contact of the two nasals. In Serpianosaurus, N. pusillus and N. peyeri, the suture between the nasals is superficially covered by the premaxilla, but the nasals continue to meet.
At the same time, the maxilla retreats from the midline. In Dactylosaurus, the maxillae broadly extend up the side of the snout, reaching well above the mid-point of the orbit and ending in a wide suture with the nasals. This broad ascending extension gradually becomes reduced to a narrow ascending process, losing a little altitude with respect to the orbit. In N. edwardsii, the ascending process is lost completely. Oddly, throughout the trajectory, the maxilla maintains control of the posterior margin of the naris.
The center of mass of the frontals and parietals consistently moves back on the skull, although the anterior margin of the frontals continues to send increasingly elaborate processes both anteriorly and posteriorly.Some of this posterior movement is at the expense of the parietal, which becomes more block-like. In fact, the parietals are wider than long in N. edwardsii. In the process, the upper temporal fenestra tends to become smaller and (except in N. peyeri) rounder. The posterior process of the postorbital consequently evolves from a long lower border for the UTF into a sharp intrusion into the body of the squamosal.
A few functional features seem clear enough. The entire system of sutures on the skull table is becoming more complex, more tightly knit and -- probably -- less flexible. The jaw muscles must have shifted considerably in the process, possibly trading efficiency and gape for speed.
Given these general trends, and the remarkably linear course of evolution in the European pachypleurosaurs, we might hazard a guess as to what was going on. At least one possibility is that pachypleurosaurs were specialist hunters trapped in a classic predator-prey arms race. Whatever habits Neusticosaurus may have had, the early pachpleurosaurs were almost certainly restricted to shallow waters. Other things being equal, prey animals in shallow waters will have less room to escape. When conditions are optimal
-- analogous to, say, senior year in high school -- selection favors those with big mouths, rapid growth, and superior dating skills. However, specialist feeders are subject to drastic population bottlenecks when, for whatever reason, prey is absent. Then -- as in the junior year of college -- selection favors individuals who can move quickly, cover more territory, and aren't too picky. In a restricted basin like the Central European Muschelkalk, a coastwise range increase isn't possible. The only way to increase range is to dive deeper and, if possible, stay down longer. Thus, the most severe evolutionary pressures, over the long run, will be for diving efficiencies: stiffer neck and body, a more hydrodynamic skull, and a stronger, more pressure-resistant skull.
Maybe. It's only a scenario. ATW070127.
Descriptions
 Dactylosaurus: Gürich 1884. D. gracilis Gürich
1884. D. schroederi Nopcsa 1928 is a junior synonym of D. gracilis [RK95].
Dactylosaurus: Gürich 1884. D. gracilis Gürich
1884. D. schroederi Nopcsa 1928 is a junior synonym of D. gracilis [RK95].
Range: Middle Triassic (Anisian?) of Europe
Phylogeny: Pachypleurosauridae:::: Anarosaurus + *.
Characters: nasals meet & suture broadly [RK95]; process of frontal separates nasals and prefrontal [1] [RK95]; UTF large, reniform [RK95]; postfrontal & postorbital enter margin of UTF [R89] [RK95]; 17 cervical vertebrae [RK95]; cervical ribs with anterior process [RK95]; dorsal centra slightly inflated [RK95]; prezygapophyses appear as deep groove in lateral view, accommodating postzygapophyses of preceding vertebra so that articulation lies in transverse plane, restricting lateral bending [R89]; dorsal ribs without pachyostosis [RK95]; pectoral girdle standard pachypleurosaur plan (see Neusticosaurus) [RK95]; coracoid proximal expansion weak [RK95]; deltopectoral crest not differentiated [RK95]; entepicondyle not identifiable [RK95]; ulna and radius relatively gracile [CG85]; three ossified carpals [RK95]; intermedium long and rectangular [RK95]; manual phalanges 2-3-3-4-3, with unguals on at least 1&2 [RK95] .
 D. gracilis Gurich 1884 is known from only the anterior end of the skeleton (below), but the forelimb, especially the ulna and radius, are much slimmer than those of Neusticosaurus. D. gracilis is the smallest known pachypleurosaur, it reached an adult degree of ossification (adult hardening of the bones) at a much smaller size than other forms. However, Rieppel & Kebang [RK95] believe the holotype is a juvenile.
D. gracilis Gurich 1884 is known from only the anterior end of the skeleton (below), but the forelimb, especially the ulna and radius, are much slimmer than those of Neusticosaurus. D. gracilis is the smallest known pachypleurosaur, it reached an adult degree of ossification (adult hardening of the bones) at a much smaller size than other forms. However, Rieppel & Kebang [RK95] believe the holotype is a juvenile.
Links: Plesiosaur Directory - Classification -
 Dactylosaurus; The systematic status of Hanosaurus hupehensis phylogeny); Fossil Room (same); Species:Dactylosaurus gracilis.
Dactylosaurus; The systematic status of Hanosaurus hupehensis phylogeny); Fossil Room (same); Species:Dactylosaurus gracilis.
Notes: [1] This is unclear. One side of Rieppel & Kebang's [RK95] reconstruction shows this feature, but the other side does not (see figure above). The specimen drawing of the cast on which the description was based shows no sign of a frontal-maxilla contact on either side.
References: Carroll & Gaskill 1985) [CG85]; Rieppel (1989) [R89]; Rieppel & Kebang 1995) [RK95]. MAK991210, ATW070113.
Anarosaurus: Dames, 1890. A. pumilio Dames 1890; A. heterodontus Rieppel & Kebang 1995; A. multidentatus Huene, 1958, has been referred to Cymatosaurus (Nothosauridae).
Range: Middle Triassic of Europe (Lower Muschelkalk -- Anisian epoch).
Phylogeny: Pachypleurosauridae ::::Dactylosaurus + *.
Characters: quite small; premaxilla broad, forming anterior half of naris [RK95]; premaxilla with slender posterior process not extending beyond posterior margin of naris [RK95]; prefrontal well-separated from postfrontal [RK95]; frontals partially fused, long & slender [RK95]; parietal foramen small [RK95]; parietal forms UTF medial margin [RK95]; parietal with tapering processes extending onto occiput [RK95]; UTF larger than in Serpianosaurus or Neusticosaurus [R89] [RK95]; postfrontal participates broadly in UTF [RK95]; postorbital ventral process with facet receiving jugal [RK95]; postorbital reaches & broadly participates in upper temporal fenestra [CG85] [R89] [RK95]; supraoccipital large, with low crest along midline [RK95]; pterygoid lateral edge with distinct ventral ridge along anteromedial & medial margins of subtemporal fossa [RK95]; retroarticular process with trough on dorsal surface [RK95] [2]; some anterior teeth with crown
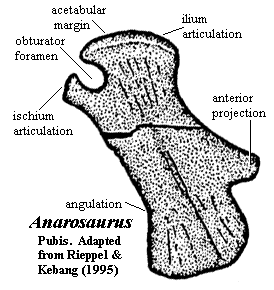 having spatulate expansion [RK95] [1]; anterior teeth elongate [R89]; 45 presacral vertebrae [RK95]; 19-20 cervical centra with ventral surface bearing paired longitudinal keels [RK95]; cervical vertebrae with pachyostosed neural arches bearing low spines [RK95]; cervical ribs distinct with anterior processes close to articular head [RK95]; large number 25-26) dorsal vertebrae [RK95]; dorsal centra slightly constricted [RK95]; ribs without pachyostosis [R89] [RK95]; gastralia with 5 elements [R89] [RK95]; 2 gastralia per vertebra [RK95]; 3 sacrals with slight proximal expansion [RK95]; humerus slightly curved, with concave medial margin, well-developed deltopectoral crest & broadly expanded distal region [RK95] humerus ectepicondylar groove present [RK95]; entepicondylar foramen well set off from distal end [RK95]; ischium with narrow acetabular region, strongly convex anterior & posterior margins, & widely expanded ventral portion with convex ventral margin [RK95]; obturator foramen open [RK95]; pubis with distinct anteriorly-directed process [3] [RK95]; femur ~30% longer than humerus (proportionately much larger than in other pachypleurosaurs) [CG85] [RK95]. ATW070113
having spatulate expansion [RK95] [1]; anterior teeth elongate [R89]; 45 presacral vertebrae [RK95]; 19-20 cervical centra with ventral surface bearing paired longitudinal keels [RK95]; cervical vertebrae with pachyostosed neural arches bearing low spines [RK95]; cervical ribs distinct with anterior processes close to articular head [RK95]; large number 25-26) dorsal vertebrae [RK95]; dorsal centra slightly constricted [RK95]; ribs without pachyostosis [R89] [RK95]; gastralia with 5 elements [R89] [RK95]; 2 gastralia per vertebra [RK95]; 3 sacrals with slight proximal expansion [RK95]; humerus slightly curved, with concave medial margin, well-developed deltopectoral crest & broadly expanded distal region [RK95] humerus ectepicondylar groove present [RK95]; entepicondylar foramen well set off from distal end [RK95]; ischium with narrow acetabular region, strongly convex anterior & posterior margins, & widely expanded ventral portion with convex ventral margin [RK95]; obturator foramen open [RK95]; pubis with distinct anteriorly-directed process [3] [RK95]; femur ~30% longer than humerus (proportionately much larger than in other pachypleurosaurs) [CG85] [RK95]. ATW070113
Comment: strangely for such a primitive form, Anarosaurus possessed fibrolamellar bone like that of pistosaurs and plesiosaurs, implying rapid growth and perhaps greater physiological and (at least when young) metabolic development. This is unlike the lamellar-zonal bone type of the stratigraphically later and morphologically more derived (advanced) Neusticosaurus which one would expect to be more advanced in this respect. It is also different to that of Nothosaurus (Klein 2010) and Ceresiosaurus (Hugi 2011). It may or may not indicate a phylogenetic relationship with primitive pistosaurs such as Cymatosaurus. MAK111103
Links: classification; Untitled Document; Species:Anarosaurus pumilio; Triassic- Netherlands;
Notes: [1] the holotype was lost or destroyed during World War II. Rieppel & Kebang [RK95] worked from deteriorating casts made 70 years earlier by Nopsca. Sensibly, they assumed that the older observations were correct, although they could not completely confirm this point with the existing material. Our illustration is probably from Zeitl, who had studied the original material.
[2] This might be the same as the facet for the posterior process of the quadrate as described for Neusticosaurus. See Jaw Notes.
[3] The same structure is found in younger specimens of Serpianosaurus [R89].
References: Carroll & Gaskill 1985) [CG85]; Rieppel (1989) [R89]; Rieppel & Kebang 1995) [RK95]. MAK991210, ATW070113.
checked ATW050109
Using this material. All material by ATW is public domain and may be freely used in any way (also any material jointly written by ATW and MAK). All material by MAK is licensed Creative Commons Attribution License Version 3.0, and may be freely used provided acknowedgement is given. All Wikipedia material is either Gnu Open Source or Creative Commons (see original Wikipedia page for details). Other graphics are copyright their respective owners