 Taxa on This Page
Taxa on This Page| Sauropterygia | ||
| The Vertebrates | Eusauropterygia |
| Vertebrates Home | Vertebrate | Vertebrate |
|
Abbreviated Dendrogram
Diapsida
├─Archosauromorpha
│
└─Lepidosauromorpha
├─Sauropterygia
│ ├─Placodontia
│ ├─Wumengosaurus
│ ├─Pachypleurosauridae
│ └─Eusauropterygia
│ ├─Corosaurus
│ └─┬─Simosaurus
│ └─┬─Nothosauridae
│ │ ├─Nothosaurus
│ │ └─┬─Lariosaurus
│ │ └─Ceresiosaurus
│ └─Plesiosauria
│ ├─Pliosauroidea
│ └─Plesiosauroidea
│ ├─Elasmosauridae
│ └─Cryptocleidoidea
└─Lepidosauriformes
├─Sphenodontia
└─Squamata
|
Contents
Overview |
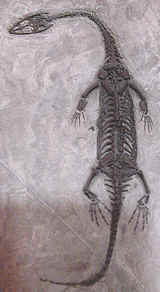
Taxa on This PageThe nothosaurids were fairly large reptiles, reaching 3 to 4 metres in length. Lariosaurus, shown at right, grew to 3 meters. They had long necks and long snouts, and probably could be thought of as a sort of cross between a crocodile and a seal. They were primarily aquatic, frequenting brackish lagoons and coastal marine environments. Most remains, such as those of Lariosaurus, have been found in Middle Triassic sediments of Switzerland and Germany (the Tethys Seaway). But the group is also known from China and elsewhere, and most probably had a world-wide tropical distribution. MAK991211.
Nothosaurs such as Nothosaurus and Ceresiosaurus have a more traditionally reptilian lamellar-zonal bone type similar to that of the pachypleurosaurs Serpianosaurusand Neusticosaurus. This is different from the fibrolamellar bone type of placodonts, pistosaurs, and some pachypleurosaurs, which implies a slower growth to maturity, and may or may not have a phylogenetic implications (Klein 2010, Hugi 2011). MAK111103
Range: Middle Triassic to Late Cretaceous
Nothosauria : Pachypleurosauridae + * : (Corosaurus + (Pistosaurus + Plesiosauria)) + (Simosaurus + Nothosauridae)
skull flattened and elongated; elongation of jaws and mandibular symphysis; orbit small relative to skull length; relative size increase in the upper temporal fenestrae; $ complex dual jaw adductor mechanism (related to rapid lateral snapping bite) to compensate for low temporal region, including separate anterior (anterodorsal force) and posterior (postero-dorsal force) adductors; loss of impedance-matching ear; strongly procumbent teeth on premaxilla and dentary; 5 premaxillary teeth; paired maxillary fangs; 5-6 sacral vertebrae; various adaptations for paraxial locomotion, i.e. rowing, rather than aguilliform motion; scapula small; . Overall trend is change from near-shore, possibly amphibious, adaptations to pelagic life.
Links: Fossil Room; Eusauropterygia after Rieppel, 1997 and Rieppel & Hagdorn, 1997.
References: Rieppel (1997). ATW020711.
Graphic: Skeleton of Nothosaurus in the Berliner Museum für Naturkunde (Berlin Museum of Natural History). Photo by Elke Wetzig, Wikipedia, GNU Free Documentation/Creative Commons Attribution
 Simosaurus: S. gaillardoti von Meyer 1842, S. guielmi von Meyer 1855
Simosaurus: S. gaillardoti von Meyer 1842, S. guielmi von Meyer 1855
Middle Triassic to Late Triassic of Europe.
Eusauropterygia :: (Nothosauridae + (Pistosaurus + Plesiosauria)) + *.
medium-sized form; head broad and flattened; snout blunt; ectopterygoid extends to cheek & secures palate [CG85]; [1]; teeth short, bluntly conical or clavate, the crown strongly striated.
Links: Fossilien aus dem Muschelkalk des Kraichgaus (German, English); Simosaurus gaillardoti (French); The Plesiosaur Site - Species.
Notes: Known only from the Upper Muschelkalk and Lettenkohle (Early to Late Ladinian epoch) of central Europe (Germany). [1] Rieppel [R89: 60-63] does a wonderful reconstruction of the jaw muscles, in a paper otherwise unrelated to Simosaurus. We could only copy the whole thing, which would be risky, as well as boring. Thus, you'll have to get it yourself.
References: Carroll & Gaskill (1985) [CG85]; Rieppel (1989) [R89]. MAK990825; ATW061217.
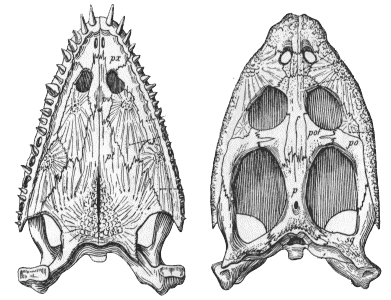
Eusauropterygia ::: (Pistosaurus + Plesiosauria) + * : Nothosaurus + (Lariosaurus + Ceresiosaurus).
M-L (0.2-4.0m); typically >2m [CG85]; long snout, small, narrow head; pterygoid vacuities closed by extension of pterygoid; extreme dorsoventral flattening of temporal region with the development of occipital flange on the squamosal and parietal, allowing posterodorsal expansion of the jaw adductor musculature; upper temporal fenestra larger than orbit [CG85]; long neck; unique pectoral girdle: essentially circular in horizontal plane with large medial opening. Scapular blades greatly reduced and extend below clavicle. Distal limb elements reduced, humerus stout. Large coracoid with posterior extension (back stroke of anterior limbs?) . Pelvic girdle somewhat similar, but more primitive.
Nothosaurus; Nothosaurus- Enchanted Learning Software; Nothosauridae; Untitled Document; some additions to Re: MOSASAURS AND PLIOSAURS; NOTHOSauria; Untitled Document; Kleiner Museumsrundgang (German).
References: Carroll & Gaskill 1985) [CG85]; Rieppel & Wild (1996). ATW061216.
 Keichousaurus: K. hui Young 1958, K. lusiensis Gonzui 1978, K. yuananensis Young 1965.
Keichousaurus: K. hui Young 1958, K. lusiensis Gonzui 1978, K. yuananensis Young 1965.
Range: Middle Triassic (Anisian) of Xingyi, Guizhou province and Yuanan, Hubei province, China.
Phylogeny: Pachypleurosauridae : Hanosaurus + ((Dactylosaurus + Anarosaurus) + (Serpianosaurus + Neusticosaurus))) + *.
Characters: postorbital enters margin of UTF [R89]; 15-30 cm; neck long; limbs sturdy; ulna massive [CG95]; five-toed feet with long digits.
Comments: The best known and most abundant of the Chinese Triassic sauropterygia, represented by many complete skeletons, This is a small pachypleurosaur with very large orbits (eye sockets) and a pointed rostrum (snout), and unusally short and broad ulna (one of the two bones of the forearm, which in plesiosaurs are incorporated into the paddle) (Lucas, 2001, p.115), unlike those of any of the European genera. Originally shown by cladistic analysis to be a basal member of a monophyletic pachypleurosauridae (Rieppel 1998, see phylogeny above), a more recent analysis shows it as a basal nothosaur (Holmes et al 2008), the implication being pachypleurosaurs as conventionally defined are a paraphyletic grade. Known to have given birth to live young (Cheng et al 2004), Sauropterygia in general were primitively vipariaous. From the abstract of the afore-cited paper: "Here we report two gravid specimens of Keichousaurus hui Young from the Middle Triassic of China. These exquisitely preserved specimens not only provide the first unequivocal evidence of reproductive mode and sexual dimorphism in sauropterygians, but also indicate that viviparity could have been expedited by the evolution of a movable pelvis in pachypleurosaurs. By extension, this has implications for the reproductive pattern of other sauropterygians and Mesozoic marine reptiles that possessed a movable pelvis."
Links: Plesiosauria com Wikipedia, Paleontology and Geology Glossary: K; Triassica - Keichousaurus hui; Dinosaur facts - Keichousaurus; GEOCLASSICS - Keichosaurus; Paleozoic.org: Gallery of Fossil Images; Fossil Reptiles & Bird Fossils For Sale; Fossil aquatic reptile, Keichousaurus from China; Peripatus -Genus Keichousaurus MAK111126
References: Carroll & Gaskill (1985) [CG85]; Rieppel (1989) [R89]. Cheng et al 2004 ATW061231. MAK991209, 111126.
 Nothosaurus: Brinckmann 1994; N. giganteus, N. mirabilis Munster. N. procerus Schröder
Nothosaurus: Brinckmann 1994; N. giganteus, N. mirabilis Munster. N. procerus Schröder
Range: Early Triassic to Late Triassic of Europe, North Africa, Russia & China.
Phylogeny: Nothosauridae : (Lariosaurus + Ceresiosaurus) + *.
Characters: ~3m; ectopterygoid extends to cheek & secures palate [CG85]; epipterygoid rather massive [CG85]; laterally flattened tail with caudal "fin"; hindlimbs longer than forelimbs;
Comments: Fossil remains of Nothosaurus are common in the Muschelkalk (Anisian and early Ladinian) of Southern Germany. Nothosaurus mirabilis Munster is the typical and best-known species, attaining a length of 3 meters. Smaller species are found in the Buntsandstein and Lettenkohle (late Ladinian).
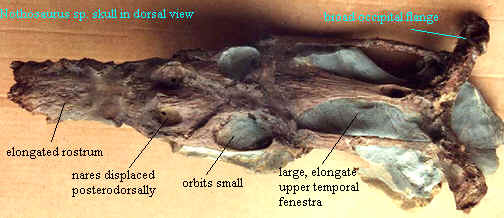
Links: Nothosaurus- Enchanted Learning Software; Nothosaurus sp. (note the elongated upper temporal fenestra, small orbits and wide occipital flange. Characteristic of secondarily aquatic reptiles, the vertebrae are disk-like and short, and the distal limb elements incompletely ossified); Untitled Document; Lecture 9 - Triassic: Moenkopi, Karroo, Ischigualasto; Fossils of the Upper Muschelkalk; Marine Reptiles: Nothosaur: Nothosaurus (test: this is a poor reconstruction. why?); Something about Pliosaurs (a completely different take on nothosaur phylogeny); Projekt \Naturkundemuseum\ (German); Nothosaurus mirabilis? Tanystropheus conspicius (Russian); Nothosaurus basic information and speculation); Nothosaurus Portuguese); NOTHOSAURUS MIRABILIS ? ??NYSTROPHEUS CONSPICIUS Russian); PANGEA Italian); Nothosaurus giganteus German -- details of minimally-prepared fossils); muschelkalk lipperland; ?????? Japanese models); Untitled Document; [노토사우루스류(Nothosaurs)] Korean); vertabrates; Winterswijk; Nothosaurus life reconstruction & a few factoids); Marine Reptiles- Nothosaur- Nothosaurus same); Lexique dinosaures- Le Nothosaurus.Fiche descriptive.Images. French: basic, but informative); Nothosaurus detailed image of a single centrum); Natural History Museum Erfurt|Nothosaurus German); Museum Wiesbaden - Naturwissenschaftliche Sammlung- Zeitreise detailed image of a ?humerus); The Plesiosaur Site - Species; Saurier fordert Techniker heraus lots of fossil images); The braincases of Simosaurus abstract).
Image: from Fossils of the Upper Muschelkalk
References: Carroll & Gaskill (1985) [CG85]. ATW061217. MAK991211.
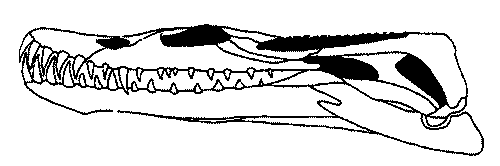 |
the skull of Nothosaurus procerus parvus Schröder length 11.5 cm Lower Muschelkalk (Anisian epoch) of Rudersdorf, near Berlin |
 Lariosaurus: L. balsamii, L. curoni, L. valcresii.
Lariosaurus: L. balsamii, L. curoni, L. valcresii.
Middle Triassic of Europe (Upper Ladinian deposits).
Phylogeny: Nothosauridae :: Ceresiosaurus + *.
Characters: cranial: skull long & low; rostrum anteriorly constricted; very long postorbital region and elongate upper temporal fenestrae, which are larger than the orbits; Parietal and squamosal rise above the occiput to form occipital crest; palate with suborbital and infratemporal vacuities; anterior teeth are very long and fang-like, procumbent and interlocking with jaws closed; pterygoid with row of teeth; axial: 17-20 cervicals, relatively short neck; cervical ribs with distinct anterior process; cervical ribs small and double-headed; 20 dorsal vertebrae; dorsal ribs very robust & pachyostotic, but degree of pachyostosis is lower in L. valceresii holotype than in; L. balsamii; 5 sacral and at least 39 caudal vertebrae; caudal ribs shorten rapidly & do not extend beyond 15th or 16th caudal; tail forming about one-third the total length; appendicular: The pectoral girdle shows the typical sauropterygian morphology: it is stout ventrally but low, and the clavicle is placed on the anteromedial surface of the dorsal blade of the scapula. The anterior limb is much stouter than the posterior one, as in other Lariosaurus species. Probably the fore-limb played a major role in swimming as suggested for nothosaurs in contrast to pachypleurosaurs. The humerus is stout, but proportionally longer and more slender in L. valceresii holotype than in L. balsami specimens. The ulna is peculiar in Lariosaurus, as it is greatly broadened while the radius is much narrower. Between radius and ulna a wide spatium interosseum is enclosed. There are five or six ossified carpals followed by rod-like metacarpals; digits short (may have had some mobility on land); only some phalanges in each digit are preserved/ossified thus impossible to give phalangeal formula for the manus in L. valceresiii; two large proximal carpals and tarsals, the distal five each supporting a digit; phalangeal formula of hand, 23443; median longitudinal and two paired lateral series of abdominal ribs (gastralia?); pelvic girdle shows wide pubis and ischium and a tiny ilium with extremely reduced iliac blade; femur slender and approximately same length as humerus in L. valceresii, but somewhat longer than the humerus in L. balsami; tibia and fibula rod- like, enclosing rather wide spatium interosseum closed distally by wide astragalus, which is larger than calcaneum; In pes, only the first two toes are completely preserved in L. valceresii holotype (second toe with four phalanges, suggesting the presence of a slight hyperphalangy in this species); hind limb longer and stouter than forelimb, but otherwise very similar; pes phalangeal formula 23454.
The humerus is stout, but proportionally longer and more slender in L. valceresii holotype than in L. balsami specimens. The ulna is peculiar in Lariosaurus, as it is greatly broadened while the radius is much narrower. Between radius and ulna a wide spatium interosseum is enclosed. There are five or six ossified carpals followed by rod-like metacarpals; digits short (may have had some mobility on land); only some phalanges in each digit are preserved/ossified thus impossible to give phalangeal formula for the manus in L. valceresiii; two large proximal carpals and tarsals, the distal five each supporting a digit; phalangeal formula of hand, 23443; median longitudinal and two paired lateral series of abdominal ribs (gastralia?); pelvic girdle shows wide pubis and ischium and a tiny ilium with extremely reduced iliac blade; femur slender and approximately same length as humerus in L. valceresii, but somewhat longer than the humerus in L. balsami; tibia and fibula rod- like, enclosing rather wide spatium interosseum closed distally by wide astragalus, which is larger than calcaneum; In pes, only the first two toes are completely preserved in L. valceresii holotype (second toe with four phalanges, suggesting the presence of a slight hyperphalangy in this species); hind limb longer and stouter than forelimb, but otherwise very similar; pes phalangeal formula 23454.
Images: Lariosaurus balsamii Curioni, specimen just under a metre in length. Middle Triassic Perledo, Lake Como, Italy. Graphic and text from Zittel, Textbook of Paleontology, vol.II, p.289. Source of life reconstruction unknown.
Note: Our profound understanding of the anatomy of this part of the Sauropterygia results from the extensive work of Dr. Olivier Rieppel of the Field Museum in Chicago. Dr. Rieppel has contributed significantly to many areas of paleontology, but the nothosaurs are his real calling. It is likely that Dr. Rieppel and Dr. Philippe Janvier are our greatest living anatomists. With luck, these skills will not be lost. It is obvious that the next century will require that the traditional anatomy be alloyed with an increasing admixture of molecular genetics (i.e. genetic signal transduction), developmental biology, and brute force genomics. However, anyone who has read the attempts of molecular biologists and genomics masters to explain evolutionary events will immediately appreciate the problem. These disciplines do not merely lack the romance and beauty of organismal biology. The practitioners of these arts have little experience working with an entire organism, much less a species or an evolutionary biota, over geologically significant amounts of time. It is, of course, exceedingly important that organismal biologists also learn the cant and incantations of their molecular and silicon-based brethren -- if nothing else, this is the one of the true paths to adequate funding. However, it is equally important that we never forget the traditional arts of observation, dissection, anatomical description, histological preparation, and the like.
Links: Lariosaurus; Nuova pagina 1 (Italian); Nuova pagina 1 (Italian); Lariosaurus balsami. ATW020629, MAK991211.
 Ceresiosaurus: C. calcagnii Peyer 1931
Ceresiosaurus: C. calcagnii Peyer 1931
Range: Middle Triassic of Europe, Lower Muschelkalk of Tessin, Switzerland. (late Anisian or early Ladinian age).
Phylogeny: Nothosauridae ::: Lariosaurus + *.
Characters: 2-4 m; ball & socket glenoid for forelimbs; long digits with hyperphalangy
Note: [1] See skeletal image. This is an odd animal. It lacks, or rather minimizes, the cranial specializations which mark the Nothosauridae. The upper temporal fenestra is not strongly elongated. The external nares are well forward, and the occipital flanges are not strong. In fact, to judge by the skull alone, it looks more like a plesiosaur. The post-cranial skeleton is equally odd, although recognizably nothosaurid, The propulsive strength has been shifted to the hindlimbs. The short and proximally wide tail suggests that the m. caudofemoralis or equivalent muscle was highly developed. The femur is short, perhaps retained within the body wall, but the lower limb is long and powerful, with extensive polyphalangy. The pelvic girdle is simplified and very solid. The Web contains numerous references stating that Ceresiosaurus was an undulatory swimmer, but this must surely be one of those sourceless scientific myths propagated by mindless cut and paste. Despite its great length, this animal was clearly engineered to be a pursuit diver, perhaps along the lines of Hesperornis, although both slower and more agile. Given a diet consisting of pachypleurosaurs
 (see Geologia Insubrica - Rivista di Scienze della Terra), this method of propulsion seems entirely reasonable. [2] Despite these differences of proportion, Rieppel has suggested that Ceresiosaurus is congeneric with Lariosaurus. See Nuova pagina 1. Detailed photographs of a new specimen of Lariosaurus (see Nuova pagina 1) make this suggestion seem more reasonable than it would appear at first glance. ATW020629. [3] The two species, C. calcagnii and C. lanzi represent a stratigraphic series, with the earlier (Anisian-Ladinian boundary) C. calcagnii being successed by the stratigraphically younger (Ladinian) C. lanzi (= Lariosaurus lanzi?). The difference between the two as regards microstructure of the long bones is similar to that of the contemporary pachypleurosaurs Serpianosaurus and Neusticosaurus with which they shared their environment, and which have been shown to constitute an anagenetic series (O'Keefe & Sander1999, the difference being that the straigraphically earlier C. calcagnii most resembles the the stratigraphically youngest pachypleurosaurid Neusticosaurus edwardsii, both having a radially orientated vascularisation pattern, whereas the younger C. lanzi is more similar to Serpianosaurus, and also to young Nothosaurus, with a longitudinally orientated pattern. The different growth pattern might be linked to a different mode of life in the restricted lagoonal basin, which ties in with the morphological data that seems to shows two different locomotion types (Hugi 2011). If Nothosaurus constitutes a more primitive or ancestral condition then, unlike the pachypleurosaurs, this is not an anagenetic series (although it can still be a phylogenetic one), because the more primitive C. lanzi preceeds the more derived (histologically speaking) C. calcagnii. MAK111103
(see Geologia Insubrica - Rivista di Scienze della Terra), this method of propulsion seems entirely reasonable. [2] Despite these differences of proportion, Rieppel has suggested that Ceresiosaurus is congeneric with Lariosaurus. See Nuova pagina 1. Detailed photographs of a new specimen of Lariosaurus (see Nuova pagina 1) make this suggestion seem more reasonable than it would appear at first glance. ATW020629. [3] The two species, C. calcagnii and C. lanzi represent a stratigraphic series, with the earlier (Anisian-Ladinian boundary) C. calcagnii being successed by the stratigraphically younger (Ladinian) C. lanzi (= Lariosaurus lanzi?). The difference between the two as regards microstructure of the long bones is similar to that of the contemporary pachypleurosaurs Serpianosaurus and Neusticosaurus with which they shared their environment, and which have been shown to constitute an anagenetic series (O'Keefe & Sander1999, the difference being that the straigraphically earlier C. calcagnii most resembles the the stratigraphically youngest pachypleurosaurid Neusticosaurus edwardsii, both having a radially orientated vascularisation pattern, whereas the younger C. lanzi is more similar to Serpianosaurus, and also to young Nothosaurus, with a longitudinally orientated pattern. The different growth pattern might be linked to a different mode of life in the restricted lagoonal basin, which ties in with the morphological data that seems to shows two different locomotion types (Hugi 2011). If Nothosaurus constitutes a more primitive or ancestral condition then, unlike the pachypleurosaurs, this is not an anagenetic series (although it can still be a phylogenetic one), because the more primitive C. lanzi preceeds the more derived (histologically speaking) C. calcagnii. MAK111103
Links: Nothosaurus- Enchanted Learning Software; Ceresiosaurus; ichthyosaur.html; Kleiner Museumsrundgang (German); Saurier aus der Mitteltrias des Monte San Giorgio (German); The Plesiosaur Site - Species; Untitled Document; Título principal aqui (Spanish); evolution4. ATW020629, MAK 991211.
Checked ATW050109
Using this material. All material by ATW is public domain and may be freely used in any way (also any material jointly written by ATW and MAK). All material by MAK is licensed Creative Commons Attribution License Version 3.0, and may be freely used provided acknowedgement is given. All Wikipedia material is either Gnu Open Source or Creative Commons (see original Wikipedia page for details). Other graphics are copyright their respective owners
