 Some Jaw Notes
Some Jaw Notes| Sauropterygia | ||
| The Vertebrates | Serpianosaurus |
| Vertebrates Home | Vertebrate | Vertebrate |
|
Abbreviated Dendrogram
Diapsida
├─Archosauromorpha
│
└─Lepidosauromorpha
├─Sauropterygia
│ ├─Placodontia
│ ├─Wumengosaurus
│ ├─Pachypleurosauridae
│ │ ├─Hanosaurus
│ │ └─┬─┬─Dactylosaurus
│ │ │ └─Anarosaurus
│ │ └─┬─Serpianosaurus
│ │ └─Neusticosaurus
│ └─┬─Nothosauridae
│ └─Plesiosauria
│ ├─Pliosauroidea
│ └─Plesiosauroidea
│ ├─Elasmosauridae
│ └─Cryptocleidoidea
└─Lepidosauriformes
├─Sphenodontia
└─Squamata
|
Contents
Overview |
Some Jaw Notes
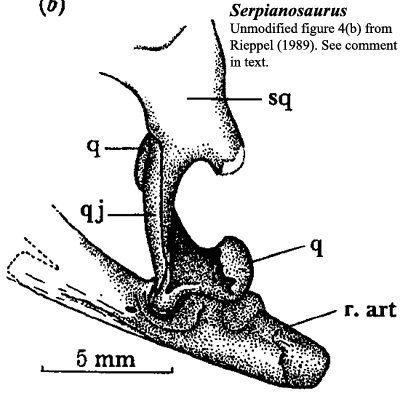
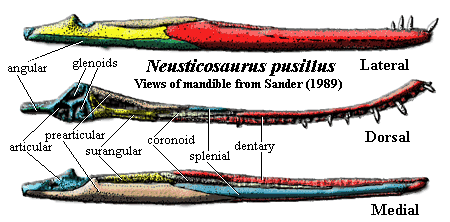
Superficial Problems: There seems to be a little confusion about the structures near the jaw articulation in the Serpianosaurus - Neusticosaurus clade. The confusion may be entirely our own. However, after carefully reviewing all three comprehensive descriptions of this area ( i.e. [CG85], [R89], and [S89]), we still don't see exactly how matters are arranged. We add this querulous note because we tried -- but failed -- to understand this tricky region enough to supply the usual colored images. Because we failed, our images of this region are inconsistent and unreliable. We supply the blow-up of an image from Rieppel's [R89] description of Serpianosaurus by way of illustrating the problem.
This image shows the jaw articulation in a sort of postero-latero-dorsal view. Most of the difficulty is related to the retroarticular process. Notice the vague labeling. Many published figures show the articular as making an appearance at the very tip of the retroarticular process, and, surely, the articular is also the surface of the lower jaw in direct contact with the quadrate. OK. Then what is the superficial bone separating the two exposures of the articular? Rieppel doesn't identify it. According to Sander [S89: fig. 4], it might be the surangular (in his fig. 4(a)), the angular (in 4(b)), or perhaps nothing (i.e. the articular exposures are not separated, as in his fig. 4(c)). Inspection of Carroll & Gaskill [CG85: fig. 14] suggests that the articular is covered distally (14e) or perhaps not (14a). If there are two separate exposures, the intervening surface may be by the surangular (14a, 14g), or perhaps the prearticular 14c).
Fitting In: For that matter, how exactly does the quadrate actually meet the articular? Unlike the first question, this one turns out to have a consistent answer. Sander [S89] provides a convincing illustration of three pits on the dorsal surface of the articular region, two sitting side-by side anteriorly, and a third just posterior to this pair. He identifies the anterior pair as receiving a double quadrate condyle, while the posterior receives the posterior process of the quadrate. This same process likely supported a tympanic membrane. [CG85] [R89]. All agree that the posterior process of the quadrate also tended to support the jaw while it was opening, and the anterior condyles carried the load during jaw closing.
Keeping a Stiff Upper Lip: This answers our question, but raises a slight additional difficulty: one that is unexplained, but probably not inexplicable. The medial part of the quadrate is also supposed to have formed a more or less rigid articulation with the recessed quadrate ramus of the pterygoid. The posterior part is supposed to have carried a tympanic membrane, as mentioned. What happened to these articulations in life? Note that water pressures would be fluctuating up and down on the tympanic membrane as a function of depth, while mechanical strains on the quadrate shifted back and forth between the three supports as a function of jaw position. The whole arrangement looks suspiciously awkward when looked at from this point of view.
Perhaps it was awkward, but we are simply overlooking a strong functional reason to maintain this complex arrangement. One of the peculiar things about pachypleurosaurs is the conflict between their anatomy and their taphonomy. Pachypleurosaurs had an inflexible quadrate with a tympanum. The usual inference from this anatomy is that they lived and hunted in very shallow seas, since their ability to dive would have been extremely limited [CG85] [R89]. However, the remains of pachypleurosaurs are actually found in conditions suggesting unusually deep, quiet waters with little postmortem transport [S89]. Sander points out, in particular, that even the gastralia and tail are often preserved in articulation. This implies that water pressure was high enough to prevent disarticulation from the usual prolonged floating and "blow-out" caused by the accumulation of decomposition gasses in the gut. Any number of odd circumstances could explain this disparity; but our favorite, among equally unsupported speculations, is that the funny-looking jaw articulation sustained a pressure-relief system for the middle ear.
 Life reconstruction of Serpianosaurus mirigiolensis. Artwork by Smokeybjb. Wikipedia, GNU Free Documentation/Creative Commons Attribution. |
Other Pressure Points: In addition to (possibly) dealing with changes in external pressures of a physical nature, the jaw articulation is also trying to tell us part of a bigger story about evolutionary pressures. As we mentioned on a previous page, this is paleohistory in the style of William Faulkner or Günter Grass. That is, the tale has lots of interesting characters and memorable vignettes; but it's hard to say what the plot is. We can't even reconstruct the exact order in which events occurred. However -- in addition to providing the reader with this gold-plated example of utterly pointless literary name-dropping -- we will list a few evolutionary issues in the pertinent Middle Triassic phylospace:
1. Shift of the main jaw articulation to the posterior process of the quadrate in other Nothosauria, but not pachypleurosaurs. Rieppel [R89] provides a very satisfactory functional explanation of this phenomenon.
2. Gradual posterior shift of the jaw articulation to a point behind the occipital condyle -- probably related to #1; except that pachpleurosaurs seem to have moved in the opposite direction.
3. Lengthening of the dentary tooth row, without a corresponding expansion of the upper jaw dentition.
4. Increase in the size of the quadratojugal and a trend towards covering the quadrate in lateral view.
5. Gradual loss of the coronoid process, but (usually) retention of the coronoid bone.
6. Increase in stiffness and consolidation of the lower jaw, the palate -- and almost everything else.
ATW070103
 Serpianosaurus: Rieppel [R89]. S. mirigiolensis Rieppel [R89]. The type was previously included in Neusticosaurus ("Pachypleurosaurus") edwardsii by Peyer 1932 and Zangerl 1935 [R89]. The type was also "informally referred" to "Phygosaurus" Arthaber 1924. [R89] argues that "Phygosaurus" is a nomen dubium, i.e., the type has been lost and the descriptions are not diagnostic to genus [R89]. S. zinae? (may be a nomen nudum).
Serpianosaurus: Rieppel [R89]. S. mirigiolensis Rieppel [R89]. The type was previously included in Neusticosaurus ("Pachypleurosaurus") edwardsii by Peyer 1932 and Zangerl 1935 [R89]. The type was also "informally referred" to "Phygosaurus" Arthaber 1924. [R89] argues that "Phygosaurus" is a nomen dubium, i.e., the type has been lost and the descriptions are not diagnostic to genus [R89]. S. zinae? (may be a nomen nudum).
Range: Middle Triassic (Anisian/Ladinian boundary -- probably restricted to earliest Ladinian) of Europe (Switzerland) [R89] [S89] [R95].
Phylogeny: Pachypleurosauridae::: Neusticosaurus + *.
Characters: medium-sized, with glenoid-acetabulum length up to 19 cm [R89];
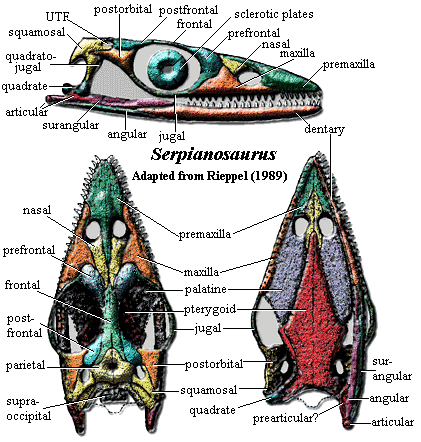 dermal skull: [1] skull relatively long [R89]; premaxilla broad, meeting maxilla broadly along suture from anteroventral naris, anteroventrally to edge of snout [R89]; anterior sides of snout straight (not convex) [R89]; maxilla long, low, forming ventral & posterior margin of naris [R89] maxilla with ascending process between naris and prefrontal [R89]; maxilla tapering posteroventral process underlying contact between anterior tip of jugal and prefrontal [R89]; strap-like nasal reaching naris, separating deep posterodorsal process of premaxilla and ascending process of maxilla [R89] frontals and premaxillae variably separate nasals, but never contact, so that nasals always have some medial suture in dorsal view [R89]; nasals without descending process [R89]; lacrimal absent [R89]; prefrontal curved, slightly inflated dorsally, forming anterior margin of orbit [R89]; prefrontal with flat process forming anteroventral margin of orbit [R89]; frontal with facet for posterodorsal process of prefrontal [R89]; frontals usually variably fused, forming dorsal orbital margins [R89]; frontal with deep posterolateral process partially separating parietal and postfrontal [R89]; frontal does not reach UTF [R89]; UTF small, bordered by squamosal, parietal, postfrontal & sometimes (narrowly) by postorbital [R89]; parietals not fused, enclosing large pineal foramen at mid-length [R89]; UTF small and sometimes triangular [R89]; transverse ridge above parietal & squamosal occipital flanges [R89]; anterior tip of jugal embedded in deep facet on posteroventral maxilla [R89]; jugal thin, curved, running between posteroventral tip of prefrontal and anteroventral tip of postorbital to form ventral and posteroventral orbit margins [R89]; postorbital triradiate [R89]; postorbital descending process receives posterior tip of jugal [R89]; postorbital posterior process deeply penetrates squamosal, but more ventrally than in Neusticosaurus [R89]; squamosal bordered medially by UTF and parietal [R89]; squamosal forms posterior part of temporal embayment [R89]; posterolateral process of squamosal covers dorsal head of quadrate in lateral view [R89]; quadratojugal relatively small, on lateral quadrate below squamosal [R89]; for articular region, see note; dorsal occiput deeply embayed with occipital flanges of squamosal and parietal [R89]; supraoccipital rectangular and "typically projects beyond the posterior margin of the parietal skull table" [2] [R89] supraoccipital with ossified ascending process (under parietals) [R89]
dermal skull: [1] skull relatively long [R89]; premaxilla broad, meeting maxilla broadly along suture from anteroventral naris, anteroventrally to edge of snout [R89]; anterior sides of snout straight (not convex) [R89]; maxilla long, low, forming ventral & posterior margin of naris [R89] maxilla with ascending process between naris and prefrontal [R89]; maxilla tapering posteroventral process underlying contact between anterior tip of jugal and prefrontal [R89]; strap-like nasal reaching naris, separating deep posterodorsal process of premaxilla and ascending process of maxilla [R89] frontals and premaxillae variably separate nasals, but never contact, so that nasals always have some medial suture in dorsal view [R89]; nasals without descending process [R89]; lacrimal absent [R89]; prefrontal curved, slightly inflated dorsally, forming anterior margin of orbit [R89]; prefrontal with flat process forming anteroventral margin of orbit [R89]; frontal with facet for posterodorsal process of prefrontal [R89]; frontals usually variably fused, forming dorsal orbital margins [R89]; frontal with deep posterolateral process partially separating parietal and postfrontal [R89]; frontal does not reach UTF [R89]; UTF small, bordered by squamosal, parietal, postfrontal & sometimes (narrowly) by postorbital [R89]; parietals not fused, enclosing large pineal foramen at mid-length [R89]; UTF small and sometimes triangular [R89]; transverse ridge above parietal & squamosal occipital flanges [R89]; anterior tip of jugal embedded in deep facet on posteroventral maxilla [R89]; jugal thin, curved, running between posteroventral tip of prefrontal and anteroventral tip of postorbital to form ventral and posteroventral orbit margins [R89]; postorbital triradiate [R89]; postorbital descending process receives posterior tip of jugal [R89]; postorbital posterior process deeply penetrates squamosal, but more ventrally than in Neusticosaurus [R89]; squamosal bordered medially by UTF and parietal [R89]; squamosal forms posterior part of temporal embayment [R89]; posterolateral process of squamosal covers dorsal head of quadrate in lateral view [R89]; quadratojugal relatively small, on lateral quadrate below squamosal [R89]; for articular region, see note; dorsal occiput deeply embayed with occipital flanges of squamosal and parietal [R89]; supraoccipital rectangular and "typically projects beyond the posterior margin of the parietal skull table" [2] [R89] supraoccipital with ossified ascending process (under parietals) [R89]
palate & braincase: anterior palate as in Neusticosaurus [3] [R89]; "the palatal shelf of the maxilla ... excludes the palatine bone from the lateral [but not posterior] margin of the internal naris [4] [R89]; vomers partially split premaxillary shelf and form medial margins of choanae [R89]; palatines very thin [R89]; pterygoids meet broadly along ventral midline, overlapping vomers & forming wide plate extending back to occiput & covering braincase [R89]; pterygoid with broad quadrate ramus meeting medial surface of quadrate [R89] quadrate ramus set off from palatal surface by curved ridge (possible origin of deep portion m. pterygoideus) [R89]; ectopterygoid probably absent [R89];
jaw& dentition: dentaries straight [R89]; dentaries weakly joined at symphysis [S89]; dentary extends to just behind a low coronoid process [R89]; angular covers ventral & lateral face of jaw [R89]; anteromedial dentary overlapped by splenial, extending almost to symphysis [R89]; articular covers ventral face of retroarticular process [R89]; but articular underlies mandibular joint [R89]; coronoid present, running anteriorly from apex of coronoid process [R89]; palatal dentition absent [R89]; teeth with thecodont implantation [R89]; 6-8 functional(??) premaxillary tooth positions [R89]; 15-16 maxillary tooth positions, with alternate replacement posteriorly [R89]; >30 dentary tooth positions, more closely spaced in middle portion [R89] [5];
 axial: long trunk (glenoid-acetabular distance 20 cm v. total length 75 cm) [CG85]; 35-38 presacral vertebrae [CG85] [R89]; centra amphicoelous [R89]; neurocentral suture present throughout [R89]; all vertebrae (except posterior caudals) bearing accessory articulation as in Neusticosaurus, with unpaired anterior medial projection from base of neural arch articulating with facet between postzygapophyses of next anterior vertebra [R89] [6]; ribs with relatively little pachyostosis [CG85]; cervicals gradually increase length & breadth in dorsal view, [R89]; cervical spines low [R89]; cervicals with short transverse process bearing diapophysis [R89]; cervical parapophysis on centrum, just posteroventral to diapophysis [R89]; ribs increase in size & curvature to anterior dorsals, becoming shorter in posterior dorsals [R89]; cervical ribs with anterior process bridging gap to (i.e., possibly articulating with) next anterior rib [R89]; transition to single-headed dorsal ribs not as abrupt as in Neusticosaurus, involving synapophysis [R89]; dorsal vertebra without parapophysis and with increasingly prominent of diapophysis on arch, which moves to anterior position on arch, with increasing breadth of transverse process [R89]; dorsal centra with smooth ventral surface [R89]; dorsal centra with slight pachyostosis across zygapophyses [R89]; prezygapophyses appear as deep groove in lateral view, accommodating postzygapophyses of preceding vertebra so that articulation lies in transverse plane, restricting lateral bending [R89]; dorsal neural spines higher posteriorly [R89]; dorsal ribs without pachyostosis [R89]; gastralia with 5 elements [R89]; 3-4 sacrals [R89]; sacral ribs not ankylosed to centra, showing broad proximal head retaining "indications of the dichocephalous condition" [R89]; sacrals with extensive transverse processes, formed by both centra and arches [R89]; most anterior caudal ribs may bend anteriorly as if associated with pelvic girdle [R89]; 47-58 caudals [R89]; caudals with no pachyostosis [R89]; anterior caudals similar to sacrals, continuing increasing height of neural spines, peaking at caudals 3-5 [R89]; transverse processes and ribs persist in tail, but become smaller and shift ventrally on centrum [R89]; caudal neural spines with increasing posterior slant [R89]; caudal ribs disappear about caudal 11 [R89]; abrupt disappearance of neural spine at caudals 15-17 [R89]; hemal arches as in Neusticosaurus, but inexplicably absent in some well-preserved specimens [R89];
axial: long trunk (glenoid-acetabular distance 20 cm v. total length 75 cm) [CG85]; 35-38 presacral vertebrae [CG85] [R89]; centra amphicoelous [R89]; neurocentral suture present throughout [R89]; all vertebrae (except posterior caudals) bearing accessory articulation as in Neusticosaurus, with unpaired anterior medial projection from base of neural arch articulating with facet between postzygapophyses of next anterior vertebra [R89] [6]; ribs with relatively little pachyostosis [CG85]; cervicals gradually increase length & breadth in dorsal view, [R89]; cervical spines low [R89]; cervicals with short transverse process bearing diapophysis [R89]; cervical parapophysis on centrum, just posteroventral to diapophysis [R89]; ribs increase in size & curvature to anterior dorsals, becoming shorter in posterior dorsals [R89]; cervical ribs with anterior process bridging gap to (i.e., possibly articulating with) next anterior rib [R89]; transition to single-headed dorsal ribs not as abrupt as in Neusticosaurus, involving synapophysis [R89]; dorsal vertebra without parapophysis and with increasingly prominent of diapophysis on arch, which moves to anterior position on arch, with increasing breadth of transverse process [R89]; dorsal centra with smooth ventral surface [R89]; dorsal centra with slight pachyostosis across zygapophyses [R89]; prezygapophyses appear as deep groove in lateral view, accommodating postzygapophyses of preceding vertebra so that articulation lies in transverse plane, restricting lateral bending [R89]; dorsal neural spines higher posteriorly [R89]; dorsal ribs without pachyostosis [R89]; gastralia with 5 elements [R89]; 3-4 sacrals [R89]; sacral ribs not ankylosed to centra, showing broad proximal head retaining "indications of the dichocephalous condition" [R89]; sacrals with extensive transverse processes, formed by both centra and arches [R89]; most anterior caudal ribs may bend anteriorly as if associated with pelvic girdle [R89]; 47-58 caudals [R89]; caudals with no pachyostosis [R89]; anterior caudals similar to sacrals, continuing increasing height of neural spines, peaking at caudals 3-5 [R89]; transverse processes and ribs persist in tail, but become smaller and shift ventrally on centrum [R89]; caudal neural spines with increasing posterior slant [R89]; caudal ribs disappear about caudal 11 [R89]; abrupt disappearance of neural spine at caudals 15-17 [R89]; hemal arches as in Neusticosaurus, but inexplicably absent in some well-preserved specimens [R89];
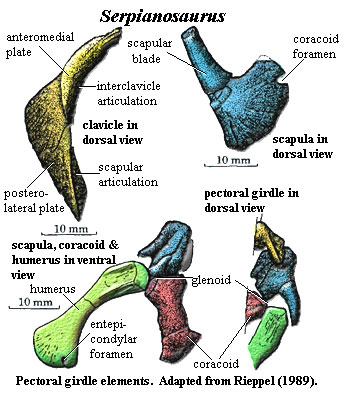
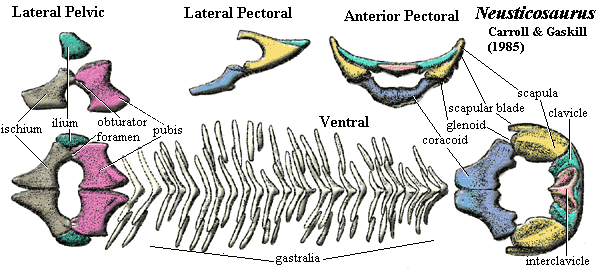
appendicular: interclavicle & clavicles form straight, solid transverse bar behind presacrals 15-18 [R89]; dermal pectoral girdle unlike Neusticosaurus in being much narrower and usually disarticulated [R89]; interclavicle entirely posteroventral and superficial to clavicle, sometimes with posteromedial extension [R89]; clavicle with anteromedial and a posterolateral plates at angle of ~120º [R89]; clavicle, posterolateral plate variably overlaps scapula [R89]; scapula with distinct ridge
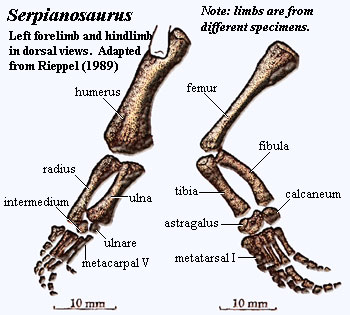 separating origins of dorsal & ventral pectoral muscles [R89]; scapula with narrow blade marked off by notch [R89]; scapular contribution to glenoid with strong ridge compared to Neusticosaurus) [R89]; medial coracoid ossified (unlike Neusticosaurus) [R89]; coracoid anterolateral corner contributes to glenoid [R89]; coracoid meets ventral scapula in straight, transverse suture [R89]; humerus weakly curved [R89]; humerus distally expanded & flattened, with smooth convex or slightly angulated articulation (separating the radial & ulnar articulations) [R89]; humerus distal articulations not extending to ventral surface (so little flexion) [R89]; ectepicondylar foramen absent [R89]; humerus with strong sexual dimorphism [R89]; radius longer and projects beyond ulna [R89]; proximal radius and entire ulna shaft with concave facing surfaces [R89]; olecranon absent, allowing epipodials to project straight out from humerus [R89]; of carpals, only ulnare & intermedium ossified [R89]; metacarpal I shorter than others, & III longest [R89]; proximal metacarpals slightly overlap [7] [R89]; phalangeal count 2-3-4-4/5-3; ilium (see Neusticosaurus image for general scheme of pelvic girdle) small & dorsally constricted = short anteroposteriorly?), broadening ventrally to contribute to acetabulum [R89]; pubis plate-like, with expanded ends, slightly angulated towards both ends [R89]; pubes & ischia contralateral bones at midline [R89]; ischium shaped very similar to pubis, with more constriction [R89]; pubis meets ischium only narrowly, to form ventral acetabulum [R89]; obturator foramen slit-like, may be completely in pubis in large specimens [R89]; femur shorter than humerus and less well-developed [R89] femoral shaft biconcave, flattened at ends, expanded proximally more than distally due to horizontal posterior extension possibly for insertion of m. iliofemoralis [R89]; fibula usually larger & longer, with much greater curvature than tibia [R89]; astragalus & calcaneum only ossified tarsals, articulating with tibia & fibula, respectively, with astragalus larger [R89]; metatarsal 1 short & broad, 4 longest [R89] phalangeal count 23454 [R89].
separating origins of dorsal & ventral pectoral muscles [R89]; scapula with narrow blade marked off by notch [R89]; scapular contribution to glenoid with strong ridge compared to Neusticosaurus) [R89]; medial coracoid ossified (unlike Neusticosaurus) [R89]; coracoid anterolateral corner contributes to glenoid [R89]; coracoid meets ventral scapula in straight, transverse suture [R89]; humerus weakly curved [R89]; humerus distally expanded & flattened, with smooth convex or slightly angulated articulation (separating the radial & ulnar articulations) [R89]; humerus distal articulations not extending to ventral surface (so little flexion) [R89]; ectepicondylar foramen absent [R89]; humerus with strong sexual dimorphism [R89]; radius longer and projects beyond ulna [R89]; proximal radius and entire ulna shaft with concave facing surfaces [R89]; olecranon absent, allowing epipodials to project straight out from humerus [R89]; of carpals, only ulnare & intermedium ossified [R89]; metacarpal I shorter than others, & III longest [R89]; proximal metacarpals slightly overlap [7] [R89]; phalangeal count 2-3-4-4/5-3; ilium (see Neusticosaurus image for general scheme of pelvic girdle) small & dorsally constricted = short anteroposteriorly?), broadening ventrally to contribute to acetabulum [R89]; pubis plate-like, with expanded ends, slightly angulated towards both ends [R89]; pubes & ischia contralateral bones at midline [R89]; ischium shaped very similar to pubis, with more constriction [R89]; pubis meets ischium only narrowly, to form ventral acetabulum [R89]; obturator foramen slit-like, may be completely in pubis in large specimens [R89]; femur shorter than humerus and less well-developed [R89] femoral shaft biconcave, flattened at ends, expanded proximally more than distally due to horizontal posterior extension possibly for insertion of m. iliofemoralis [R89]; fibula usually larger & longer, with much greater curvature than tibia [R89]; astragalus & calcaneum only ossified tarsals, articulating with tibia & fibula, respectively, with astragalus larger [R89]; metatarsal 1 short & broad, 4 longest [R89] phalangeal count 23454 [R89].
Links: The Plesiosaur Directory - Images; Saurier aus der Mitteltrias des Monte San Giorgio
Notes:
[1] For general morphology, see images of Neusticosaurus skull and jaws.
[2] Nope. We have no idea what this means, either.
[3] In comparison with Neusticosaurus, the palate is very poorly known. The vomers appear to be considerably smaller in Serpianosaurus so that the anterior palate is dominated by the palatal shelf of the premaxillae. The braincase of Serpianosaurus is completely unknown.
[4] This is not at all clear in either the reconstruction or specimen drawings. Consequently, our images do not reflect this relationship.
[5] As Rieppel [R89] notes, more teeth on both jaws than Neusticosaurus.
[6] Rieppel [R89] does not find the second, more ventral, accessory articulation found in some Neusticosaurus.
[7] The implication is probably that they spread like a fan.
References: Carroll & Gaskill 1985) [CG85]; Rieppel (1989) [R89]; Rieppel (1995) [R95]; Sander (1989) [S89]. MAK991211, ATW061231, rev'd ATW080328.
revised ATW080328
Using this material. All material by ATW is public domain and may be freely used in any way (also any material jointly written by ATW and MAK). All material by MAK is licensed Creative Commons Attribution License Version 3.0, and may be freely used provided acknowedgement is given. All Wikipedia material is either Gnu Open Source or Creative Commons (see original Wikipedia page for details). Other graphics are copyright their respective owners
{kind=link}
{kind=link}
{kind=link}