Theropoda: Basal Tetanurae : The Megalosaurs
Taxa on this Page
- Afrovenator X
- Dubreuillosaurus X
- Duriavenator X
- Eustreptospondylus X
- Marshosaurus X
- Megalosaurinae X
- Megalosaurus X
- Piatnitzkysaurus X
- Poekilopleuron X
- Tetanurae
- Torvosaurus X
- Xuanhanosaurus X
Basal Tetanurae, aka the Megalosaurs
As with Pelycosauria and Synapsida, Thecodontia and Archosauria, and Podokesauridae and Neotheropoda, we are once again at one of those phylogenetic and evolutionary nodes where evolutionary systematics gives one term to refer to a stem group consisting of similar forms from which many other evolutionary lineages diverge, while cladistics gives a totally different name to refer to that exact same evolutionary node and the exact same taxa, but also including all the descendents, regardless of how distinct they are. In this instance, the Megalosauridae are a family (and grade) of large but otherwise unexceptional Jurassic theropods that replaced the large podokesaurs (dilophosaurs) and were in turn replaced by their descendents the allosaurs in the late Jurassic. The Tetanurae are a clade of theropods that include not only all the megalosaurs but everything that evolved from them, or from their descendents or their descendents descemndents, including tyrannosaurs, ostrich dinosaurs, segnosaurs, proto-birds, modern birds of every sort, and many other wondrous creatures, many of which bare not the slightest resemblance to their big bodied, short but stocky fore-armed, ancestors. What they do have in common though are certain shared traits (or synapomorphies) by which the clade is defined.
The situation is complicated however by the fact that - unlike the Pelycosauria vis a vis the Synapsida, or the Thecodonts re the Archosaurs - not everyone agrees that megalosaurs are an ancestral group; some argue that they are a sister clade of the more bird-like avetheropods. This depends on the type of cladogram that results from a phylogenetic analysis; some analyses show the megalosaurs are an artifical (in cladistic, not evolutionary, terms) group, others as a monophyletic clade. We have followed occam's razor in arguing the former, the reason being that megalosaurs are both morphologically and stratigraphically intermediate between Early Jurassic dilophosaurs (advanced podokesaurs) and Mid to Late Jurassic avetheropods; if they aren't the ancestors then where are they? In other words, the apparent monophyly of the megalosaurs is - as with the ceratosaurs - an artifact of cladistic analysis, due to the problems such as reversals, homoplasies, and so on MAK120222
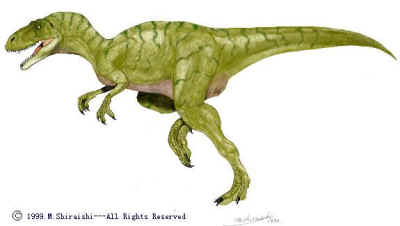 The Megalosaurs presumably evolved from a dilophosaur- like ancestor during the earlier Jurassic, becoming the dominant large preditor by the middle Jurassic, before being in turn supplanted by the Allosaurs. Cretaceous forms were specialised semi-aquatic fish-eaters, mostly confined to Gondwanaland, and sometimes attaining very large size and a Dimetrodon-like sail on the back. They seem to have died out some time during the earlier late Cretaceous, along with their contemporaries the giant carcharodontosaurs. The terms Megalosaur, Megalosauridae, etc were unbtil recently rarely used, as firstly the original or type species of Megalosaurus is known from only very fragmentary material, and secondly Megalosaurs are a paraphyletic group, defined mostly by shared primitive features, and hence not considered valid in the current, cladistic, paradigm, although a phylogenetic analysis is emerging that sheds new light on this fascinating but little known period of theropod history.
The Megalosaurs presumably evolved from a dilophosaur- like ancestor during the earlier Jurassic, becoming the dominant large preditor by the middle Jurassic, before being in turn supplanted by the Allosaurs. Cretaceous forms were specialised semi-aquatic fish-eaters, mostly confined to Gondwanaland, and sometimes attaining very large size and a Dimetrodon-like sail on the back. They seem to have died out some time during the earlier late Cretaceous, along with their contemporaries the giant carcharodontosaurs. The terms Megalosaur, Megalosauridae, etc were unbtil recently rarely used, as firstly the original or type species of Megalosaurus is known from only very fragmentary material, and secondly Megalosaurs are a paraphyletic group, defined mostly by shared primitive features, and hence not considered valid in the current, cladistic, paradigm, although a phylogenetic analysis is emerging that sheds new light on this fascinating but little known period of theropod history.
The Tetanurae ("stiff tails") consist of a number of parallel lineages of theropods, all of which seemed to have evolved increasingly bird-like features. For example, their rib-cages indicate they had a sophisticated air-sac-ventilated lung system, which exists today only in birds. Such an advanced respiratory system would have been accompanied by an advanced circulatory system (even the ectothermic crocodiles (the nearest living dinosaur-ancestors after birds) have an efficient four-chambered heart, like mammals and birds, rather than the inefficient three-chambered reptile heart). All this indicates a high metabolic rate; like birds, advanced theropods were certainly endothermic (warm-blooded).
The clade Tetanurae includes both birds and the most famous of classic theropods. All Tetanurans lack the fourth digit of the hand, have all their maxilliary (upper jaw) teeth in front of their eyes, have a strap-like scapula (shoulder blade), and various other anatomical characteristics, which indicate that all tetanurans evolved from a single common ancestor. Tetanurans are generally subdivided into two clades, the Carnosauria and the Coelurosauria.
Megalosaurs represent a sort of intermediate group that does not fit in neatly with either the Ceratosauria or the Tetanurae, although nowadays the tendency is to consider at least some megalosaurs (or Torvosauroidea or Spinosauroidea, to give alternative names) as the most primitive (basal or underived) members of the Tetanurae "stiff tails"), a clade that includes both birds and the advanced theropods. Megalosaurs were previously included with the Allosaurs and Tyrannosaurs in the polyphyletic (artificial) taxon "Carnosauria." The old-style definition of Carnosaurs is actually an ecotype rather than a true evolutionary group, designating any large (around 200 kg or more in weight) carnivorous theropods, posessing large skulls, short necks, and small forearms. MAK010506 revised 120221
Descriptions
Tetanurae: Definition: birds > Ceratosaurus. Padian et al. 1999)
Range: From Early Jurassic.
Phylogeny: Averostra : Ceratosauria + * : Xuanhanosaurus + (Megalosaurinae + (Eustreptospondylus + Spinosauridae) + ((Piatnitzkysaurus + Marshosaurus) + (Afrovenator + Dubreuillosaurus) + Avetheropoda))
Characters: $ Increased pneumaticity of skull; nasals narrow & elongate [H+01]; $ maxillary fenestrae present; $ maxillary teeth only anterior to orbit(?) or to antorbital fenestrae(?); procoelous dorsal vertebrae [H+01]; stiffened tails (reduced importance of tail in walking?); scapula straplike [H+01]; coracoid tapers posteriorly [H+01]; $(?) metacarpals 1&2 broadly in contact [H+01]; $(?) manus with 3 digits; manus elongate [H+01]; $(?) obturator notch on ilium; $ fibula reduced and "clasped" by tibia; $ anterior horizontal groove on astragalar condyles. ATW
Comments: As indicated, the name of the clade refers to the development of a stiffened tail; members of the clade are also characterised by the loss of an external fourth digit on the hand, though some basal members did retain the fourth metacarpal (Holtz 1998). Most basal members of the Tetanurae are large species, the 'carnosaurs' of older authors. Within the Tetanurae, a well supported clade, variously called Avetheropoda and Neotetanurae, unites the Coelurosauria with the Allosauroidea. (adopted from CKT)
Sereno's (1999) diagnosis of the Tetanurae seems to include features more appropriate to Avetheropoda and Maniraptora. ATW
Links: The Tetanuran Theropods: Diverse carnivorous dinosaurs - UCMP; Tetanurae - Wikipedia; Tetanurae - Thescelosaurus!, includes listing of every species; Mikko's Phylogeny a dendrogram and references; Variety of Life: Tetanurae - another dendrogram and references; Tetanury (Tetanurae) bazalne detailed description in Polish, with oodles of cladograms at the end; Megalosauroidea part of Mickey Mortimer's theropod pages, not just every every taxon but every museum specimen listed down to the smallest detail; the most comprehensive (if rather technical) coverage you will find anywhere. Checked MAK120223
References: Hutt et al. (2001) [H+01].
Torvosauroidea : ( = Spinosauroidea, Megalosauroidea)
Synonyms: Torvosauridae Jensen, 1985, Eustreptospondylidae Paul, 1988, Streptospondylidae Kurzanov, 1989
Phylogenetic definitions: assuming a monophyletic Megalosauridae: (1) any dinosaur that shares a common ancestor with Poekilopleuron valesdunensis (= Dubreuillosaurus), Torvosaurus, and Afrovenator (Allain 2002) (2) all dinosaurs more closely related to Megalosaurus than to Spinosaurus, Allosaurus, or modern birds (represented by Passer domesticus). Holtz et al 2004 (3) (as Torvosauridae) same definition as Holtz but replacing Megalosaurus with Torvosaurus (Sereno 2005, Taxon Search) (via Wikipedia).
Range: Middle to Late Jurassic of Eur, NthAm, & Afr
Characters: In essence, all Tetanurae other than Avetheropods. May be para- or polyphyletic. Known material is scrappy and includes both highly derived Spinosaurs as well as relatively general Allosaur-like Torvosaurids. Elongated rostral ramus of maxilla; pinched rostral ramus of lacrimal; $ large sickle claw on manus 1.
Comments: A distinction should be made between a monophyletic, cladistic Megalosauridae and a paraphyletic, similarity-based, Megalosauridae; the one being far more restrictive than the other (corresponding perhaps to only a single linnaean subfamily or tribe). The former can be defined on gross morphology alone, and represnts one of those stem bubbles at the base of a romerogram. the latter is very hard to determine, and, as with Ceratosauria and Coelophysoidea (= Podokesauridae), there is the problem of homoplasies and reversals which hide the original morphology-based phylogenetic signal. So, cladistically, this taxon is monophyletic according to Allain 2002, Holtz et al. 2004 and Benson, 2010, but paraphyletic according to Smith et al 2007, Smith et al 2008, Sadlier et al 2008 and Xu et al 2009. Of course in either definition there would still be a monophyletic Megalosauridae, assuming Megalosaurus bucklandii isn't the literal ancestor of later theropods (and here remember that unlike evolutionary systematics, cladistics does not accept actual ancestors, only hypothetical ones), but the monophyletic clade would be greatly restricted, perhaps to only a few species. A paraphyletic definition makes sense not only morphologically (by allowing a continuous sequence of evolution forms) but also stratigraphically by eliminating the annoying ghost lineage regarding the absense of Jurassic spinosaurs.
This group includes Megalosaurus, Torvosaurus, Poikilopleuron, and possibly also Edmarka, Erectopus, and Xuanhanosaurus. These were large animals ( upto 9 or more meters long), with short, stout arms. Paul Sereno rejected the use of the name Megalosauridae, due to the fragmentary nature of Megalosaurus, and used the name Torvosauridae (based on the well known late Jurassic form Torvosaurus. Hence Torvosauroidea and so on. This was a shame I feel, because Megalosaurus has a nice historical heritage. The English species Megalosaurus bucklandii was the very first dinosaur to be described, some years before Richard Owen coined the term Dinosauria in 1842. Unfortunately, the name Megalosaurus, like Plesiosaurus, became something of a taxonomic waste basket, and there is some doubt now among paleontologists whether it even is a valid genus. That is, although the remains are of a large primitive theropod dinosaur of "megalosaur" (or torvosaur) relationships, it is not possible to identify them more closely. At present there are three species still included. At times Megalosauroidea and Torvosauroidea were replaced by Spinosauroidea. More recently Megalosaurus, Megalosaridae, and Megalosauroidea have made a comeback, and the old historical trem now seems to be the accepted term for this clade of large early primitive theropods (e.g. Benson et al, 2010). At the same time, some of these taxa, such as Eustreptospondylus and Piatnitzkysaurus, are sometimes considered to have allosaurian or basal avetheropodan attributes (Paul 1988, With all of these basal tetanurans, the precise phylogenetic placement, and even whether they are related to each other in the first place, is not clear MAK010506, revised 120222
Xuanhanosaurus qilixiaensis Dong, 1984
Horizon: Lower Shaximiao Formation of China (Bathonian-Callovian)
Phylogeny: Tetanurae/Megalosauroidea/Megalosauridae : (Megalosaurinae + (Eustreptospondylus + Spinosauridae) + ((Piatnitzkysaurus + Marshosaurus) + (Afrovenator + Dubreuillosaurus) + Avetheropoda)) + *
Size: length about 4.5 meters, weight 250 kg (Paul 2010)
Comments: Known from a partial skeleton, this robust, moderately sized theropod is remarkable in that it has a large heavily-built forelimb, which led to the idea it was quadrupedal, although it is more likely that it used its arms to grab prey. It is rarely include din cladograms, but when it is it tends to appear near or at the bottom of the tree, as a stem tetanuran, as in Holtz et al 2004. Benson et al, 2010 places it near Marshosaurus and Piatnitzkysaurus, although in vuiew of the highly endemic nature of Chinese Jurassic dinosaur faunas this may not be likely. Cau & Maganacu's cladogram (unpublished data, online) place it beneath the megalosaurs. MAK120222
Megalosaurinae Huxley, 1869 : Megalosaurus, Torvosaurus, ?Poekilopleuron.
Definition: same as Megalosauridae sensu Holtz et al., 2004; Megalosaurus bucklandii > Spinosaurus aegyptiacus, Allosaurus fragilis, Passer domesticus
Synonyms: Torvosauridae Jensen, 1985
Range: Middle to Late Jurassic of Eur & NthAm
Highly speculative Phylogeny: Tetanurae/Megalosauroidea/Megalosauridae : Xuanhanosaurus + ((Eustreptospondylus + Spinosauridae) + ((Piatnitzkysaurus + Marshosaurus) + (Afrovenator + Dubreuillosaurus) + Avetheropoda) + * : Duriavenator + ( Megalosaurus + Torvosaurus)))
Comments: Considered monophyletic by Sereno et al. (1998), Allain 2002, Holtz et al. 2004 and Benson, 2010, paraphyletic taxon or evolutionary grade by Holtz 2000, Smith et al 2007, Sadlier et al 2008, Xu et al 2009. Status of Poekilopleuron is uncertain, it may be a sinraptorid (Benson et al, 2010). Other taxa like Afrovenator and Eustreptospondylus may be associated with Avetheropoda and Spinosaurids MAK120224
Duriavenator hesperis
Synonyms: Megalosaurus hesperis Waldman, 1974
Horizon: Upper Inferior Oolite of Dorset, England (late Bajocian)
Phylogeny: Megalosaurinae : (Megalosaurus + Poekilopleuron?) + Torvosaurus) + *
Size: about 5 meters? weight about 300 kg (Paul 1988)
Comments: Known only from teeth and snout fragments, this is another of those scrappy Jurassic theropods. From the little that is known of it this early, medium-sized animal seems to be quite similar to M. bucklandii. (Paul 1988). It was recently redescribed by Roger Benson, who found that several features show it to be distinct from the lectotype dentary of Megalosaurus (Benson, 2008a); He therefore erected a new genus, Duriavenator (combining the Latin name of Dorset, Duria, with Latin for "hunter", venator) Wikipedia. Cladistic analysis places it at the base of the Megalosaurine (Torvosaurine) line (Benson, 2010, Benson et al, 2010). With Magnosaurus, it is one of the oldest-known tetanurans. MAK120221
Megalosaurus bucklandii Meyer, 1832
Horizon: Stonesfield Slate, Inferior Oolite, Great Oolite, Forest Marble, and Coralline Oolite of England, unamed unit of Indr, France (Bathonian-?early Oxfordian)
Phylogeny: Megalosaurinae : Duriavenator + (Torvosaurus + (Poekilopleuron? + *))
Size: length about 7 to 7.3 meters, weight about 1.1 tonnes (Paul 1988)
Comments: The first dinosaur to be named, this specimen is known from fragmentary remains. The original specimen was a dentary, but many other elements have been referred to it, including teeth and postcrania . Few dinosaurs have gone through so many changes in intrepretation, from being considered the thigh bone of a biblical giant, to being likened to a set of testicles (Scrotum humanum, a name that technically has priority of Megalosaurus bucklandii, although fortunately for theropod nomenclature it has never been applied), to a crocodile-headed Victorian antedilevian quadraped, to the familiar bipedal theropod, as shown below.
 From Wikipedia: Megalosaurus may have been the first dinosaur to be described in the scientific literature. Part of a bone was recovered from a limestone quarry at Cornwell near Chipping Norton, Oxfordshire, England in 1676. The fragment was sent to Robert Plot, Professor of Chemistry at the University of Oxford and first curator of the Ashmolean Museum, who published a description in his Natural History of Oxfordshire in 1676. He correctly identified the bone as the lower extremity of the femur of a large animal and he recognized that it was too large to belong to any known species. He therefore concluded it to be the thigh bone of a biblical giant, such as those mentioned in the Bible. The bone has since been lost but the illustration is detailed enough that some have since identified it as that of Megalosaurus. The Cornwell bone was described again by Richard Brookes in 1763. He jokingly named it Scrotum humanum. Image: Cover of Robert Plot's Natural History of Oxfordshire, 1677 (right), and illustration of a fossilized lower extremity of a Megalosaurus femur (left) taken from that book. Public domain, via Wikipedia. |
 Modified from Wikipedia: In 1852, Benjamin Waterhouse Hawkins was commissioned to build a model of Megalosaurus for the exhibition of dinosaurs at the Crystal Palace in Sydenham, where it remains to this day. Huge and visually impressive even by today's standards, Megalosaurus became one of the park's three 'mascot dinosaurs' along with the Iguanodon and (less so) Ichthyosaurus. Yet it is arguably the most inaccurate of all of Hawkins's models, with a huge crocodile-like head and walking on all fours. The hump on the back of the sculpture in Crystal Palace and other restorations from the 1800s was based on the material now referred to as Becklespinax. Image:
Megalosaurus at the Crystal Palace, London. Photo by C. G. P. Grey; Creative Commons Attribution, via Wikipedia. |
 It was not until the middle of the nineteenth century, when more complete remains of theropod dinosaurs began to be discovered in North America, that a more accurate picture of these animals as long tailed bipeds developed. Megalosaurus was soon supplanted by its better known American counterparts, Allosaurus and Tyrannosaurus. The iconic image of Megalosaurus however, and one that exerted a strong influence on the present author (MAK) during his childhood dinosaur studies, is the representation by the now sadly little known paleo artist Neave Parker. Considering his work predates the dinosaur renaissance of the 70s and 80s, his Megalosaurus, shown taking long strides with a horizontal back and elevated tail, is remarkably modern in posture, the only criticism being the head should be held above the body with the neck in the classic archosaurian S-shaped posture. Image: Love in the Time of Chasmosaurus - Vintage Dinosaur Art: Dinosaurs - British Museum (Natural History) MAK120224 |
For a long time the genus Megalosaurus was something of waste basket or taxon for poorly known mostly Jurassic theropods; it is currently associated only with the type species, M. bucklandii. This was a rather big animal, clearly the top preditor of its day. Cladistic analysis sometimes (Holtz et al. 2004, Benson et al, 2010) places it close to Torvosaurus, although it has also been placed separately (Sadlier et al 2008, Mortimer) MAK010506
Poekilopleuron bucklandii Eudes-Deslongchamps, 1838
Synonyms: Poecilopleuron (l. c.) AC02, Poecilopleurum (l. c.) AC02, Poicilopleuron (l. c.) AC02, Poikilopleuron (l. c.) AC02]; Poekilopleuron bucklandii Eudes-Deslongchamps 1838 [=Megalosaurus poikilopleuron Huene 1926, Poecilopleuron bucklandi (l. c.), Poecilopleurum bucklandi (l. c.), Poekilopleuron bucklandi (l. c.), Poicilopleuron bucklandi (l. c.), Poikilopleuron bucklandi (l. c.)] AC02 - CKT110613
Horizon: Calcaire de Caen, Calvados, France (early Bathonian)
Phylogeny: Megalosaurinae : Duriavenator + (Megalosaurus + (Poekilopleuron? + *))
Size: length about 7 meters, weight about 1 tonne (Paul 1988)
Comments: Poekilopleuron is to French paleontology what Megalosaurus is to English, and quickly became a similar taxonomic wastebasket. Add to that the fact that it is almost impossible to spell, leading to innumerable synonyms. The fate of the original specimen was rather unfortunate. Apparently fairly complete when found, a lot of the material was destroyed before collection, and the rest during WWII. The taxon has since had to be studied on the basis of cast replicas. Subsequent discoveries of theropod remains from France may belong to this species (Paul 1988). The most distinctive part of this animal are its forelimbs. Quoting Wikipedia: "Their length, about 60 cm, was a sign of this theropod's more original build. Unlike later Theropoda, whose forelimbs tended toward reduction in length in proportion to the animals' size, Poekilopleuron's were long and, by implication, potent. The length mostly resided in the elongated but powerfully muscled humerus. The antebrachia (forearms) were markedly short and robust, a characteristic shared with Poekilopleuron's slightly later and considerably larger American cousin Torvosaurus." Its phylogenetic position is uncertain. It may be related to or identical to M. bucklandii, or some material assigned to the latter may belong to this species. (Paul 2010). Considered part of a monophyletic Megalosauridae close to Eustreptospondylus and Dubreuillosaurus by Allain 2002, and sister taxon of Megalosaurus + Torvosaurus by Holtz et al. 2004; conversely placed in the Sinraptoridae by Benson et al, 2010 this may be yet another exmple of mosaic evolution, an intermediate form between megalosaurids and sinraptorids MAK010506, revised 120224
References: APS03, AC02 CKT110613
Torvosaurus tanneri Galton and Jensen 1979
Synonyms: Edmarka rex Bakker, Kralis, Siegwarth, and Filla, 1992
Horizon: Middle Morrison Formation of Colorado, Wyoming, & Utah (early Tithonian)
Phylogeny: Megalosaurinae : Duriavenator + (Megalosaurus + Poekilopleuron?) + * )
Size: length about 9 meters weight about 2 tonnes (Paul 1988, 2010)
Comments: Known from a partial forelimb, pelvis, and skull, this was among the largest of the Morrison preditors. This animal was larger and more heavily built than Allosaurus. Yet it has a number of primitive features that indicate relationships with typical Middle Jurassic theropods like Gasosaurus and Poekilopleuron. The absense of a calcaneal notch, already present in the much earlier Eustreptospondylus, indicates that, like Ceratosaurus, this animal was a sort of "living fossil", continuing to survive and flourish alongside much more specialised and advanced theropods MAK010506. This species has also been reported from Portugal (Mateus et al 2006
Reference: Paul 1988
Links: Torvosauroidea; Re- Torvosauroidea was [R- Torvosaurus & Giganotosaurus];
Eustreptospondylus oxoniensis Walker, 1964
Synonyms: Megalosaurus cuvieri, Streptospondylus cuvieri ; Monotypal rank: Eustreptospondylidae Paul, 1988, Streptospondylidae Kurzanov, 1989
Horizon: Middle Oxford Clay, England (late Callovian), also NW France - Late Callovian/Early Oxfordian (for Streptospondylus altdorfensis)
Phylogeny: Tetanurae/Megalosauroidea/Megalosauridae : Xuanhanosaurus + (Megalosaurinae + ((Piatnitzkysaurus + Marshosaurus) + (Afrovenator + Dubreuillosaurus) + Avetheropoda) + (Spinosauridae + * ))
Size: length 4.63 meters (adult would be larger), weight 218 kg (juvenile) (Paul 1988)
Comments: known from a partial skull and skeleton of juvenile/subadult, this is among the better known of the middle Jurassic theropods, and until the discovery of Baronyx the most complete theropod skeleton from England. According to Greg Paul (1988), this animal is more or less intermediate between Megalosaurus and more advanced forms like Allosaurus, Cladistic analysis hawever found Eustreptospondylus to be a fairly typical megalosaur (Sereno et al. (1998), Allain 2002), albeit in a different clade to Megalosaurus and/or Torvosaurus (Holtz et al. 2004, Benson, 2010) , a basal megalosaurid (Ortega et al 2010), or in a paraphyletic Megalosauria (Holtz 2000).
Similarily, the family Eustreptospondylidae was coined by Gregory Paul to include a number of advanced megalosaur and primitive allosaur like animals. All have the allosaur-like flexible ball-and-socket neck articulation, but in other respects they are too primitive to be considered allosaurs. More recent cladistic analysis shows the family as Paul defined it (which included Eustreptospondylus, Piatnitzkysaurus, Gasosaurus, and Marshosaurus to be an artificial assemblage, and it would seem that their allosaur-like chracteristics are either convergences or shared primitive features. One analysis still finds a clade centered on Eustreptospondylus, which could be call Eustreptospondylinae, as a subset of the Megalosauridae Benson, 2010.
Others find Eustreptospondylus to be a sister taxon/stem spinosaur or place it with the spinosaurs (Smith et al 2007, Sadlier et al 2008, Smith et al 2008, Xu et al 2009, Cau, et al 2011), so perhaps it's a megalosaurid evolving towards the spinosaurid condition. As no Jurassic spinosaurids are known (apart from a few referred teeth, which could be anything), this would make Eustreptospondylus a nice non-missing link. In view of the gradational nature of evolution, it would not be surprising if clades like Megalosauroidea and Megalosauridae go the way of the old clades Ceratosauria and Coelophysoidea, to be replaced by a stepwise series of smaller lineages. In any case, this shows that there is no sharp dividing line between megalosaurids, spinosaurids, and allosaurs, but rather evolutionary gradations.
The very poorly known Streptospondylus is often placed near Eustreptospondylus (Allain 2001, Smith et al 2007, Smith et al 2008) alone or, in addition to that taxon, the even more poorly known Magnosaurus (Benson et al, 2010) . Streptospondylus altdorfensis Meyer, 1832 is a contemporary French species known from assorted postcrania (Allain & Pereda Suberbiola 2003). Even more than most 19th century Jurassic European theropods known from scrappy material, this animal is a taxonomic nightmare, see the Wikipedia page for more. This may be the same species as E. oxoniensis in which case the earlier name has priority Greg Paul (2010)). Or it may be the same genus but a slightly different species. For now we have decided to retain Eustreptospondylus MAK010506 revised 120222
Piatnitzkysaurus floresi Bonaparte, 1979
Horizon: Cerro Condor Formation of Argentina (Callovian/Oxfordian)
Phylogeny: Tetanurae/Megalosauroidea/Megalosauridae : Xuanhanosaurus + (Megalosaurinae + (Eustreptospondylus + Spinosauridae) + ((Afrovenator + Dubreuillosaurus) + Avetheropoda + (Marshosaurus + *)))
Size: length about 4.5 meters, weight 275 kg (Paul 1988)
Comments: Known from two partial skulls and a partial skeleton this moderately sized theropod and originally described as a primitive allosaur (Bonaparte, 1979, Bonaparte, 1986). It was a lightly built medium-sized bipedal carnivore with robust forearms. The discovery of Piatnitzkysaurus was important in that it was the first South American theropod known from fairly complete remains and still remains the only well-known Jurassic South American theropod Despite its fairly complete status, it shares with most other middle Jurassic theropods a problematic phylogentic placement. In other words, there is little agreement as to where it belongs on the theropod evolutionary tree. It has has variously been considered a basal tetanuran (Rauhut 2003, Smith et al 2007, Xu et al 2009, Rauhut et al 2010), united with other medium-sized taxa such as Piatnitzkysaurus and Marshosaurus in a distinct lineage of basal tetanurans or basal megalosauroidea (Benson, 2010, Benson et al, 2010), as a basal monophyletic megalosaur (Holtz et al 2004, as a primitive (Sadlier et al 2008) or an advanced (Holtz 2000) taxon of a paraphyletic megalosaur sequence, an allosaur-like (transitional) "eustreptospondyline" megalosaur (Paul 1988), a basal avetheropod (Paul 2010, Cau, et al 2011), or a primitive allosaur (Chure, 2000). Since this pretty much covers every lineage of early Mid Jurassic theropod other than proto-tyrannosaur, we can sum up by saying that no one seems to know what it is. Of course, that is nothing unusual as far as megalosaurs go. The problem with placement of "transitional" mid Jureassic theropods like Piatnitzkysaurus and Monolophosaurus (which in very similar fashion has likewise been located everywhere from basal tetanuran to stem allosaur, depending on tree topology) is that in parsimony analysis of atomistic character traits, it is not possible to determine if a taxon is very primitive and has not yet acquired apomorphies (unique traits) of more derived clades, or is very advanced and has lost those same synapamorphies (shared traits) through reversal. If either or both Piatnitzkysaurus and Monolophosaurus are stem tetaneura or stem-megalosauroidea, this means that allosaurian traits have evolved repeatedly by convergence. if they are stem-avetheropods it means that some very primitive traits are retained even in advanced intermediary forms, a common phenomenon known as mosaic evolution. Either explanation is equally plausible. For now, we have placed these taxa as stem avetheropods, although future studies may well result in revising this phylogeny.
Assuming a stem-avetheropod interepretation, Greg Paul's observation (Paul 1988) that this Jurassic South American animal is more advanced than the South American abelisaurs that lived later, in the Cretaceous period is pertinant, although we now know that highly derived allosaurs (megaraptorans) continued alongside abelisaurs even to the end of the Createcous.
Condorraptor currumili Rauhut 2005 seems to be a closely related form (Xu et al 2009, Benson et al, 2010). MAK010506 revised 120227
Marshosaurus bicentesimus Madsen, 1976
Horizon: Morrison formation of Utah and ?Colorado (Late Kimmeridgian )
Phylogeny: Tetanurae/Megalosauroidea/Megalosauridae : Xuanhanosaurus + (Megalosaurinae + (Eustreptospondylus + Spinosauridae) + ((Afrovenator + Dubreuillosaurus) + Avetheropoda + (Piatnitzkysaurus + *)))
Size: length about 4.5 meters , weight about 225 kg (Paul 1988)
Comments: Known from a partial skeleton, this is a poorly known animal of uncertain relationships, it may be a primitive allosaur, or alternatively a megalosaur. It is intermediate in both size and evolutionary development between the contemporary small but advanced coelurosaurs and large but primitive ceratosaurs, but also the same size as the earlier Gasosaurus and Piatnitzkysaurus. The pelvis alone contains features reminiscent of coelophysoids, coelurids, sinraptorids, and allosaurids. The forearms seem to have been quite short (a megalosaurid feature). Benson et al, 2010 places it as a basal tetanuran close to Piatnitzkysaurus MAK010506
Afrovenator abakensis Sereno, Wilson, Larsson, Dutheil, and Sues, 1994
Horizon: Tiouraren Formation (Bathonian-Oxfordian) of Niger
Phylogeny: Tetanurae/Megalosauroidea/Megalosauridae : Xuanhanosaurus + (Megalosaurinae + (Eustreptospondylus + Spinosauridae) + ((Piatnitzkysaurus + Marshosaurus) + Avetheropoda + (Dubreuillosaurus + *)))
Size: length 8 to 9 meters, weight 1 or 2 tonnes (Paul 2010)
Comments: Known from the majority of the skull and a partial skeleton, this is one of of the better known types of dinosaur, and in fact the most complete skeleton of a Mesozoic large theropod ever found in Africa (most dinosaurs are only known from a few scraps of bone). This was a lightly built animal, with long legs implying a pursuit predator. the head is long and low, with proportionally large teeth (Paul 2010) this respectable-sized animal seems similiar to Eustreptospondylus (Allain 2002, Holtz et al. 2004), although it has also been placed as basal avetheropod (Holtz 2000). A recent cladistic analysis place it in a more basal position in the Megalosaurine (Torvosaurine) line ( Xu et al 2009. Benson, 2010, Benson et al, 2010). It has also been placed near the avetheropoda (Smith et al 2007, Sadlier et al 2008), and these two latter findings may be complementary; i,e, Afrovenator may reprsent a transitional form between megalosaurids/ines and avetheropods. In any case there seems to be little difference between all the species at the base of the tetanuran family tree. Originally believed to be of Hauterivian (Early Cretaceous) age (Sereno et al., 1994), a sort of "living fossil" alongside its more advanced cousins, the Tiouraren Formation in which Afrovenator has been found is now considered Mid Jurassic in age (Rauhut and Lopez-Arbarello, 2009). Closley related to Dubreuillosaurus, indicating that these animals ranged widely MAK010506 021221
Dubreuillosaurus valesdunensis (Allain, 2002) (or Afrovenator valesdunensis
Range: Mid Bathonian of France
Phylogeny: Tetanurae/Megalosauroidea/Megalosauridae : Xuanhanosaurus + (Megalosaurinae + (Eustreptospondylus + Spinosauridae) + (Avetheropoda + (Piatnitzkysaurus + Marshosaurus) + (Afrovenator + *)))
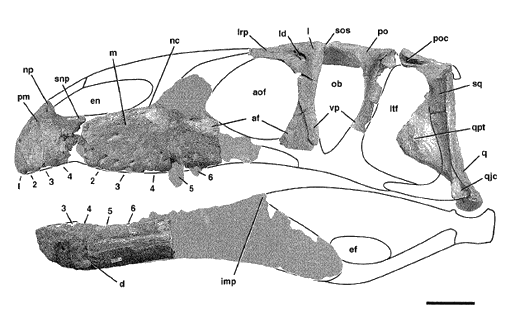
Comments: Known from an excellent skull and fragmentary postcrania (the result of the skeleton being blown to pieces when the quarry where the fossil was found was reopened). Originally assigned to the genus Poekilopleuron (Allain & Pereda Suberbiola 2003), this megalosaur had an unusually low and long skull, with the length of the skull being three times the height. Cladistic analysis consistently places it close to the contemporary genus Afrovenator (Holtz et al. 2004, Smith et al 2007, Xu et al 2009, Benson, 2010, Benson et al, 2010), perhaps it should be placed in the same genus, although it would make the name Afrovenator a seem bit incongruous for a European species. MAK120221
Links: Thescelosaurus, Wikipedia, Theropod Database (Mickey Mortimer - places this taxon in a monophyletic Eustreptospondylinae) MAK120224
Image Artwork by Nobu Tamura - Wikipedia Gnu/3.0 - by-



{kind=link}