Theropoda: Avetheropoda
Cryolophosaurus, Monolophosaurus, & Sinraptor
Taxa on this Page
- Allosauroidea X
- Avetheropoda
- Gasosaurus X
- Lourinhanosaurus X
- Metriacanthosaurus X
- Monolophosaurus X
- Sinraptor/Yangchuanosaurus dongi X
- Sinraptoridae X
- Yangchuanosaurus shangyouensis X
Carnosauria

Carnosauria was originally used as a general term for all of the big theropods, including the elusive and quite possibly non-existent, Megalosaurus, the Snark of dinosaurs. In the 1920's Huene suggested that, despite its position as the biggest and baddest theropod, Tyrannosaurus looked a lot more as if it belonged with the "small" theropods, the Coelurosauria. However, this was the Jurassic of paleontology, and most dinosaur paleontologists were lumbering, slow-moving, low-metabolism forms. With the advent of cladistic adaptations in the 1980's, dinosaur paleontologists became faster, hungrier and more predatory. The more derived forms began dismembering virtually defenseless nineteenth century taxa with lightning-fast computers, developing, in the process, enlarged manual phalanges from entering data. Others began hunting in packs, with alpha professors leading vicious gangs of slavering, pencil-wielding graduate students through the Mesozoic halls of their museums in search of prey.
Just in this manner was Carnosauria decimated. At first, only the oldest and weakest members, like Megalosaurus, were picked off. But the blood lust of the new paleontology knew no bounds. In just a few short years, all that was left of Carnosauria were Allosaurus and a few meager, nondescript bones at the bottom of the largest and rustiest drawers of the most obscure museums.
Recently, however, intensive conservation efforts by Tom Holtz and others have resulted in a remarkable worldwide resurgence. With the discovery of Giganotosaurus and related forms, Carnosauria is once again the home of the world's largest terrestrial predators. Cryolophosaurus and other, as yet undescribed, species from Gondwanaland could lead to a renaissance of the Carnosauria. Even so, the threat of extinction remains very real. Paul Sereno, backed by the awesome commercial might of the National Geographic Society, has sworn to exterminate the Carnosauria and crush its quivering remains into Allosauroidea, leaving nothing but coelurosaurs and a random miscellany of basal "neotetanurines" i.e. avetheropods).
Citizens! This is the taxon that made Victorian Englishmen gasp in horror at the Crystal Gardens in 1854. This is the clade that made movie critics shudder at "Carnosaur." This is the group that even made paleontologists scream at the publication in a kids' book of "Elvisaurus." Save the Carnosaurs!
Sinraptoridae
Something about carnosaurs seems to draw the best from people. Some of the most extraordinary monographs in paleontology were inspired by the giant Jurassic killers, and Currie & Zhao (1993) is one of them. It is well-written, copiously illustrated, reflects careful comparison with related forms, and contains some useful reflections on the functional significance of some of the more important morphological findings. It is everything a monograph should be.
It is important to be clear on the many strong points of this article, because I am going to take issue with its one weak point. Many readers will of the article will not have the several hours it takes to work through the entire piece. In fact, perhaps the majority will only read the family and species diagnosis at the beginning, and one or two anatomical points of special interest to them.
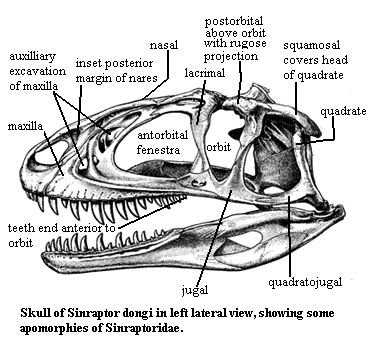
The problem is that there appear to be some significant errors or ambiguities in the family diagnosis. These are contained in a long list of characters which unite sinraptorids with allosaurids as follows:
"upper quadratojugal prong of jugal shorter than jugal prong."
Sinraptor has three prongs on the quadratojugal process of the jugal. the external (i.e. lateral) prongs are roughly the same size. It is the medial prong that is long. Dorsoventrally, this is neither the upper or lower prong, but falls between the lateral prongs. The medial prong is likely homologous to the lower prong of two in, e.g., Allosaurus, but the description as given is either confusing or erroneous.
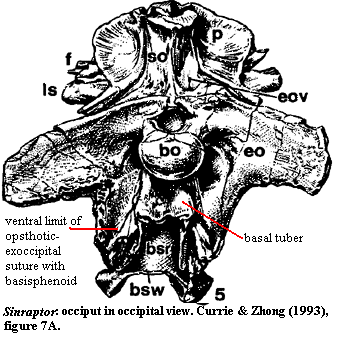
"braincase box has opened up posteriorly between tubera, which are no longer pedunculate."
The actual conformation of the occiput is shown in Currie & Zhao's Figure 7A, which is reproduced at right. The braincase box, as I understand the term which is not used elsewhere in the text), does not open up posteriorly, and there is nothing between the basal tubera which are, in fact, at least slightly pedunculate. Perhaps the authors are referring to some other tubera. Perhaps "posterior" was not the correct indication. Perhaps I have misunderstood the text. However, at a minimum, the diagnosis is unclear.
"oval muscular attachment on basioccipital."
Nothing at all is said about this in the text.
"posteroventral limit of the exoccipital-opisthotic contacts the basisphenoid but is separated from the basal tubera by a notch."
This may be correct, but it is not clear from the actual description, which reads:
Sinraptor is unusual in that the exoccipitals extend significantly more ventrally than the basal tubera. In most theropods, including ... allosaurids ... the ventral limit of the exoccipital-opisthotic along the basioccipital and basisphenoid suture is either at the same level or is dorsal to the level of the basal tubera. A notch separates the basal tuber from the more anteroventral extension of the exoccipital-basisphenoid suture as in allosaurids.
Id. at 2049 (citations omitted)
So, the notch is common to sinraptorids and allosaurids. However, the entire morphology of the occiput here is quite distinct. The similarity exists but is not likely to be an indication of common ancestry.
"Tenth presacral vertebra in process of incorporation into dorsal, rather than cervical region."
What could "in process of incorporation" mean? What the text actually indicates is that the 10th presacral vertebra is a cervical vertebra, although it bears some similarities with the tenth presacral in Allosaurus, which is a dorsal vertebra. (Id. at 2057 & 2065).
"triangular neurapophysis."
What is meant here is apparently "triangular neurapophysis on the axis-atlas complex." (see id. at 2055).
The really interesting and peculiar thing about sinraptorids may be the way in which the head was held. As Currie & Zhong note: "The shapes and alignment of the axial intercentrum and centrum served to bring the neck underneath the occipital condyle to support the skull in a manner that is unique amongst Jurassic theropods, with only Monolophosaurus, Y[angchuanensis] magnus and an undescribed theropod showing a tendency towards doing the same thing." (Id. at 2057, citations omitted.)
This posture is reflected in life restorations of Sinraptor and Allosaurus by Shiraishi Mineo. The head of Sinraptor is held higher. The anterior part of the back is arched to compensate (as also noted by Currie & Zhao). The difference in carriage may well be responsible for a number of differences between Allosaurus and Sinraptor, such as the anteroventral extension of the opisthotic-exoccipital, the use of the 10th presacral vertebra as a cervical, rotation of the atlas intercentrum, and apparently greater emphasis on stiffening the back with hyposphene-hypantrum articulations.
To speculate further on function, one might envision Sinraptor as a slower, but more maneuverable, version of Allosaurus. Sinraptor was more anteroposteriorly compact and laterally rigid. It might well have been able to shift its weight rapidly from side to side, as suggested by Currie & Zhao's notes on locomotion discussed in the main entry. The raised position of the head combined with rapid shifts in its center of gravity would also allow Sinraptor to attack over a wider angle compared to the antero-posterior axis of its body when the attack began. Running in this position would be awkward, but Sinraptor would have the advantage in an ambush attack. Perhaps this is also the reason for the fact that Sinraptor has an even more completely pneumatized skull than Allosaurus. Allosaurus would rely on the lightness and speed of its entire body. Sinraptorwould rely more on the speed of its head. This is, perhaps, a parallel development to the changes in posture which led to the upright carriage of birds.
Descriptions
(= Neotetanurae = Allosauria = Allosauroidea of some authors)
Early Jurassic-Late Cretaceous.
U-shaped premaxillary symphysis; tapering posterior of coracoid; proximal half of metacarpal I closely appressed to metacarpal II; pubic boot; loss of obturator foramen; cnemial process originates on lateral surface (of tibia).
Monolophosaurus, Allosaurus,, etc.
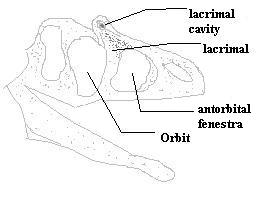
Large, long, narrow skull; $ enlarged nares; $ nasal participates in antorbital fossa; pre-orbital horns, commonly based in lacrimal; antorbital fenestra is expanded upwards; $ large cavity in lacrimal at antero-dorsal margin of orbit (possible gland of unknown function); skull commonly crested; large orbits; cheek flat, vertical, at sharp angle to skull roof [CC00]; quadrate with broad quadratojugal flange; quadrate short, with head at mid-orbit level; basioccipital excluded from basal tubera (but Sinraptor?); $ basipterygoid process very short; basi- parasphenoid formed by delicate-looking thin webs of bone; opisthocoelous vertebrae; pubic boot longer anteriorly than posteriorly; pubic boot triangular with apex posterior in ventral view; femur larger than tibia.
With the exclusion of Tyrannosauroidea (now considered a giant Coelurosaur lineage), only the Allosaurids and their relatives make up the Carnosauria. The name "Allosauria" was coined by Greg Paul in his superb book Predatory Dinosaurs of the World and seems to have caught on, although "Carnosauria" is still sometimes used. In any case, these were big to enormous bipedal terrestrial preditors, that for some one hundred million years or so were the dominant form of life on land.
The stem-based definition of Carnosauria is more inclusive than the node-based definition of Allosauroidea (definitions from Carnosauria (in Polish)):
- Carnosauria (Huene, 1920): the largest clade that includes Allosaurus fragilis but not Passer domesticus (modified from Padian and Hutchinson, 1997)
- Allosauroidea (Marsh, 1878 vide Currie and Zhao, 1994) the smallest clade that includes both Allosaurus fragilis & Sinraptor dongi (Holtz, Molnar and Currie, 2004; modified from Padian and Hutchinson, 1997)
With new phylogenies reshufling basal tetanuran and avetheropodan taxa, some of which were only recently described to begin with, it has become hard to know if a primitive theropod is a basal Allosauroid, a non-Allosauroid (i.e. stem-taxon) Carnosaur, a basal Megalosauroid, or a basal Tetanuran! Monolophosaurus for example has been considered all of these at some point or another. With the main debates in early theropod evolution being now about whether a taxon belongs in a monophyletic Megalosauroidea or paraphyletic basal Tetanura, or either of the latter or Avetheropoda, or vs Avetheropoda, most cladograms now have a simple node that is not distinguished further for re Allosauroidea/Carnosauria.
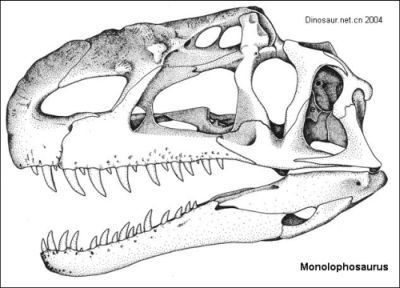 |
| Monolophosaurus skull
|
Junggar Basin, Xinjiang, China
Dorsal surface of nasal convex and rugose; facial horns present [CC00]; dorsal surface of frontals convex [CC00]; some caudal vertebrae with hyposphenes; supraacetabular shelf continuous with lateral margin of pubic peduncle.
standard total length of about 5.5 meters (Paul 2010)
Known from a partial skeleton and compressed skull, Monolophosaurus is a medium-sized early theropod distinguished by a single very large midline ridge-like head crest running from its nose to the rear of the skull. Beyond that, there is nothing particularily distinctive about it, and, as with its megalosaur cousins, exactly where it belongs on the theropod evolutionary tree is a matter of some controversy. The situation here seems to be very similar to that of Piatnitzkysaurus, another mid Jurassic theropod that has had the misfortunate of being bounced all aroudn the theropod family tree by rival phylogenetic analyses.
Originally, things were pretty straightforward. Holtz 2000,Holtz et al 2004, Novas et al 2005, and Corria & Currie 2006 are among the references that considered it an early allosauroid, which places it here among the avetheropods. Pretty soon however things started to go stemward. Smith et al. (2007), in a radical study using Cryolophosaurus (until then considered an allosaur) reinterpreted the positions of many taxa, and demolished the old clades Ceratosauria and Coelophysoidea, and found Monolophosaurus to be as a basal avetheropod. Similarily, Rauhut 2003 had previously found this species to be part of a paraphyletic megalosauria leading to allosaurs. Worse was to come with Benson, 2008b, in a paper on European megalosaurs, noting various primitive features of the skull and skeleton suggest that Monolophosaurus is actually a basal megalosauroid. This position was also recovered in a cladogram by Cau, et al 2011 in a paper on carcharodontosaurs. The implication here would be that characters previously considered apomorphies of the Allosauroidea are plesiomorphic (shared primitive traits, not unique to allosaurs) (Zhao et al. 2010 and Brusatte et al. 2010). Mickey Mortimor mega-matrix analysis finds it to be either a sister to Avetheropoda or a basal megalosauroid. As against placement as a basal tetanuran, a cladogram in the recent a paper on on the megalosaur Eustreptospondylus (Sadlier et al 2008) and another of the advanced allosaur Acrocanthosaurus atokensis (Eddy & Clarke (2011)) both recover Monolophosaurus as a basal allosauroid. This brings us full circle, and we are back where we started after oceans of ink and reams of computer print-outs. It is pretty clear here as elsewhere that the choice of taxa and traits keyed into to the analysis would determine the outcome of the cladogram. It may be that a monophyletic Allosauroidea has to be replaced by any number of parallel megalosaurian lineages. Or this may simply be an example of mosaic evolution, a common feature in transitional taxa, so that early allosaurs or avetheropods were megalosaur-like, primitive basal megalosaur-like, in many ways. Or that Mid Jurassic theropod evolution is such a jumble that a lot more material is required before any even half complete phylogeny can emerge. For now, we've gone with the mosaic evolution option and retained Monolophosaurus as a basla carnosaur, although Smith et al. (2007)'s similar interpertation of this species as a basal avetheropod may be equally valid
Incidentally, Monolophosaurus wasn't the only theropod with a mono loph. A similar but even more exaggerated feature is found in the contemporary basal tyrannosaur Guanlong wucaii. If Monolophosaurus is a basal tetanuran then the two are not related and such characters would arise through convergence ( Brusatte et al. 2010). If Monolophosaurus is a primitive avetheropod (whether stem-avetheropod or stem-carnosaur) then the two would only have fairly recently diverged, and these sort of crests may well be a common shared characteristic among basal avetheropods. The appareance of paired crests among ancestral neotheropods (podokesaurs) and horns and ridges among ceratosaurs and abelisaurs illustrates the repeated occurance of exotic head structures among theropods, doubtless for intra-specific display.
Monolophosaurus has been related to another taxon, Chuandongocoelurus (Benson, 2008b), known only from a single medium-sized partial skeleton, although the association isn't very strong (see Theropod database for comments), and was later considered a more basal form (Ortega et al 2010). If they are related they would constitute an endemic theropod clade (to which we could give the informal name Monolophosauridae (Bakker 1997), so far known from the Middle Jurassic of Asia (Zhao et al. 2010). But this may however be yet another example of convergence. Also, because no skull is known for Chuandongocoelurus, comparisons with Monolophosaurus and other taxa are made more problematic.
Usually 7-10m long, laterally compressed. Long, narrow skull; flange-shaped lacrimal process on palatine; $ articular with pendant(?) medial process; $ participation of nasal in antorbital fossa; antorbital fenestra expanded dorsally; maxillary fenestra expanded dorsally to form hole in lacrimal (probably plesiomorphic for carnosaurs); pneumatic excavation of ascending ramus of maxilla (same character, described differently); lacrimal recess, single opening present (another aspect of the same character state); narial prominences comprised of paired ridges along lateral edges of nasals; postorbital prominences present; orbits large; lacrimal horns and crests common; quadrate articultion projects well ventral of ventral surface of maxilla; horizontal shelf on lateral surface of surangular, rostroventral to mandibule condyle, prominent and pendant; caudal dorsal neural spines oriented cranially; three fingers per hand; manus digits 2 & 3 roughly equal with 2 slightly longer; pubic boot triangular (apex caudal) in ventral view, acute angle between shaft and caudal portion of boot; $ femur usually larger than tibia; 2 distal tarsals (III & IV).
Allosauroidea proper is now considered a monophyletic clade divided into three similiarily monophyletic families: the Sinraptoridae, the Allosauridae, and the Carcharodontosauridae.
Greg Paul considers the small late Jurassic Ornitholestes and the earlier and larger Proceratosaurus to be also Allosaurs. This has not caught on, and current understanding classifies Ornitholestes as a basal (i.e. primitive/ancestral) coelurosaur, while Proceratosaurus has recently been identified as a basal tyrannosaur.
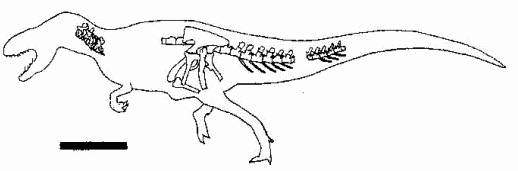 |
| Type of Lourinhanosaurus antunesi. Scale bar 0.5 m.
|
late Kimmeridgian/early Tithonian
Amoreira-Porto Novo of Lourinha, Portugal
Known from a fragmentary skeleton, this medium-sized theropod may be a sinraptorid (Mateus (1998), Benson, 2008, 2010) an unspecified carnosaur (Holtz et al 2004), an allosaur (Paul 2010), , or a distinct non-avetheropod tetaneran clade that also includes Streptospondylus (Mickey Mortimer). In any case, it represents a more primitive form than Allosaurus and Acrocanthosaurus (Mateus (1998)). If it is a sinraptorid it shows the lineage was established in Europe as wlel as China. Of course, Sinraptorid attributes may simply be common primtive characteristics, as well as some specialised traits that were lost in more advanced forms. This means that whilst Lourinhanosaurus may be a sinraptorid grade theropod, it still may or may not be necessarily a sinraptorid in the strict sense
Sinraptor, Yangchuanosaurus, Metriacanthosaurus?, Poekilopleuron?
$ at least 2 accessory pneumatic excavations of maxilla; complex, variable network of pnumatic fossa associated with maxilla and nasals; maxilla rugose only above tooth row; maxilla longer than tall [CC00]; external nares with marked inset of posterior margin; antorbital sinus extends into lacrimal, nasal & jugal; ventral process of lacrimal with 2 convexities and process for suborbital ligament (marking ventral extent of eyeball); lacrimal does not contact maxilla; dorsolateral horn of lacrimal low; nasal very long, flat, and smooth; smooth internasal articulation (nasals may have been able to move upward and away from each other); nasal with pneumatic openings; nasal provides groove for ridge on internal process of maxilla (may allow some outward rotation of maxilla); tight sutures of nasal with lacrimal and frontal; small triangular prefrontal; frontal flat dorsally; no sagittal crest; strong nuchal crest; ventral end of postorbital above ventral margin of orbit; bulbous, rugose anterior projection on post-orbital; complex, layered jugal-maxilla articulation; $ posterior quadratojugal) process of jugal has 3 prongs with medial prong much longer (2 equal prongs in other carnosaurs; note that only the two lateral prongs can be seen in the figure); jugal & quadratojugal do not invade lateral temporal fenestra [CC00]; $ squamosal flange covers head of quadrate in lateral view; parietal forms obtuse angle with occiput [CC00]; paroccipital process broad & shallow, turning ventrally at lateral ends, with distal posterior projection [CC00]; large crista prootica on prootic; downturned paroccipital process; basioccipital participates in small basal tubera (but see Sereno,1997); paroccipital process turns downwards slightly; slight or "incipient" separation of exit for opthalmic branch of Vth cranial nerve; vomers fused anteriorly only; vomers contact pterygoids; symphysis of lower jaw probably flexible; dentary makes sliding contact with surangular and angular; elongate articular with retroarticular process; 4 pairs of premaxillary teeth; 15 maxillary teeth [CC00]; last maxillary tooth anterior to orbit; teeth with "tooth plates (modified cingula?) on lingual side; tooth plates not fused; triangular neurapophysis on atlas-axis complex; 10 opisthocoelous cervical vertebrae, but 10th is similar to allosaurid dorsal; neck held below skull in unique fashion; dorsal vertebrae amphiplatyan; back arched in thoracic region; hyposphene - hypantrum articulation on all dorsal vertebrae; 5 sacral vertebrae; some caudal vertebrae with hyposphenes; pectoral girdle largely unknown; sternum smaller than in Allosaurus; manus digit 1 relatively slight; vestigial manus 4 present; wide supraacetabular shelf continuous with lateral margin of pubic peduncle (see figure); brevis shelf narrow; pubes meet in symphysis along distal half of length; moderately expanded pubic boot; well-developed obturator process; strongly developed lesser & 4th trochanters; leg swung out (abducted) when swung forward (protracted) & hips shifted over opposite leg; lower leg generally same as in Allosaurus; astrgalus tightly linked to tibia; astragalus has extensive contact with fibula; ascending process of astragalus low.
These are the most primitive allosauroids, although even at this early stage they were equipped with elaborate crests. While several European theropods have been placed here, it is not sure if they belong here, as many known only from quite inadequate material. Sinraptorids proper are known from China (hence sinraptorids), but that is more likely because Chinese dinosaur fossils are the best representatives of the middle Jurassic, rather than the group itself being limited to that region. However, it is still possible that Eastern China was cut off from the rest of Laurasia, as a distinctive fauna (Shunosaurine sauropods, Beinotheroides tritylodontids, and brachyopoid labyrinthodonts, seem to be indiginous to that region.
(= Yangchuanosaurus dongi)
Junggar Basin, Xinjiang, China
Known from complete skulls and the majority of a skeleton, this was a typical large theropod. It's cranial ornamentation is rather modest. S. hepingensis is a closely reated species, or perhaps a synonym (see Thescelosaurus! website). If Yangchuanosaurus and Sinraptor are co-generic, than the former name has priority, and this species becomes Yangchuanosaurus dongi (Paul 2010)
The holotype of Sinraptor dongi is not fully mature. The remains bear unhealed bite marks (possibly related to the animal's death), as well as healed broken ribs.
Y. magnus Dong, Zhou, and Zhang, 1983
The largest carnivore of its time, this three and a half tonne preditor dominated its enviroment. The cranial ornamentation involves dual ridges above the snout and eyes. It also has a low fin fin, down its back, again, possibly for display purposes. Greg Paul (1988) considered this species similiar enough to go under Metriacanthosaurus. It is very similar to S. dongi
Megalosaurus parkeri Huene, 1926
early or middle Oxfordian
From the little that is known of this animal (a few postcranial elements: some vertebrae, a femur) it appears similar to the Sinraptorids, where it is usually placed (Paul 1988, Benson, 2010). The two also lived at more or less the same time. The presence of fin-like ridge along back (as indicated by the tall-spined vertebrae) meant that for a long time this animal was considered an ancestral spinosaurid (Steele, 1970). As Justin Tweet (Thescelosaurus! website) notes, following a brief fifteen minutes of fame in the 1980s (thanks to Greg Paul's Predatory Dinos of the World, associating it with Yangchuanosaurus, or rather vice-versa) and 1990s (because of Paul's influence the name showed up in the movie version of Jurassic Park), this taxon sank back into obscurity.
Because the more recent name Sinraptoridae (Currie & Zhao 1994), is used instead of the first published name, Metriacanthosauridae (Paul 1988), it has been suggested the latter should take precedence. But because it was not accompanied by a formal description, it cannot be considered a scientific valid under formal rules of nomenclature. In any case it is no longer used, not even by Greg Paul himself, we have not tried to revive it. Perhaps this is understandable given the scrappy nature of the eponymous genus. We've placed Metriacanthosaurus next to the sinraptorids, but it may well turn out to be a totally different lineage of early avetheropod. If neither Metriacanthosaurus or Lourinhanosaurus are sinraptorids, then the latter taxon is endemic to China, home of many unique Jurassic dinosaurs
checked ATW050619, this page MAK120304

{kind=link}