Theropoda: Ceratosauria
Taxa on this Page
- Abelisauroidea X
- Ceratosauria X
- Ceratosauridae X
- Ceratosaurus dentisulcatus X
- Ceratosaurus nasicornis X
- Deltadromeus X
- Elaphrosaurus X
- "Limusauridae" X
- Limusaurus X
- Spinostropheus X
Abelisaurus, artwork courtesy of Frank DeNota
Ceratosauria - the other theropods
For more than one and a half centuries, dinosaur discoveries were characterised by predominantly northern (Laurasian) faunas, reflecting the fact that science (including paleontology) was centered in countries like Britian, France, Germany, and the United States, and that fossils from these countries (and their expeditions) would feature strongly and determine the way we thought about the evolution of life on Earth. So in the case of the Mesozoic, we think of the Late Triassic Keuper (Germany) and Chinle (SW USA) formations, and animals like Plateosaurus and Ceolophysis, the Early Jurassic the Liassic Scelidosaurus of England and Middle Jurassic Megalosaurus and Cetiosaurus of the same country and Poekilopleuron and Omosaurus/Lexovisaurus of France, late Jurassic the Morrison (central West USA) and Tendaguru (East Africa - German expedition) with their spectacular megafauna, the Early Createceous Wealden of England (with its classic trio of Megalosaurus, Iguanodon, and Hylaeosaurus), and the abundant latest Certaceous dinosaurs of Alberta, Montana, and Wyoming, the rich fauna of tyrannosaurs, ostrich dinosaurs, horned dinosaurs, duckbills, and the rest. When fossils from other parts of the world were discovered, they were slotted into northern taxa, so in the case of theropods, Elaphrosaurus was a coelurosaur (then an ornithomimosaur), Genyodectes a megalosaur, Indosuchus a megalosaur or an allosaur
In the late 20th century, dinosaurology underwent multiple shifts of focus. John Ostrom and his student Bob Bakker initiated the dinosaur renaissance and the realisation that dinosaurs were active animals closely related to birds, in China spectacular discoveries were made of feathered dinosaurs, supporting Ostrom and Bakker's hypothesis, and in South America (mostly Argentina) spectacular discovceries wer made not only of the earliest dinosaurs at the base of the family tree, but Cretaceous faunas very different to those of the ornithischian dominated north, including meat-eating dinosaurs that did not fit in to the normal model of theropod evolution.
When beginning in the mid 90s these Southern (Gondwanan) theropods were analysed via cladistic methods, it was realised that they constituted a large group totally disntinct from the familiar northern faunas. These are the Ceratosauria - the other theropods.
The Ceratosauria are a lineage of persistantly primitive theropods that suddenly appear in the fossil record around the later Middle/earlier Late Jurassic mark and continue right through to the end of the Cretaceous, co-existing with the standard theropods (or tetaneura). Phylogenetically, they are defined as including all species more closely related to Ceratosaurus than to birds. As well as larger forms such as Ceratosaurus and the Abelisauridae, the ceratosaurs also included smaller forms such as the primitive Limusaurs and the advanced Noasaurids. Although Ceratosaurus itself came from the Jurassic of North America, these animals had dissapeared from the northern continents by the early Cretaceous. By the Middle to Late Cretaceous this formerly minor lineage had come to dominated the medium to large carnivore niche not only throughout in Gondwana (with representatives known from South America, Africa, India, and possibly Australia), but in Europe as well. It is not known why these more primitive animals were able to supplant their more advanced, birdlike, and successful tetanuran cousins (equivalent to the if Cretaceous marsupials had to supplant Cenozoic mammals); indeed, this is a good example of how misleading terms like "primitive" and advanced" can be, in terms of evolutionary success. MAK120210
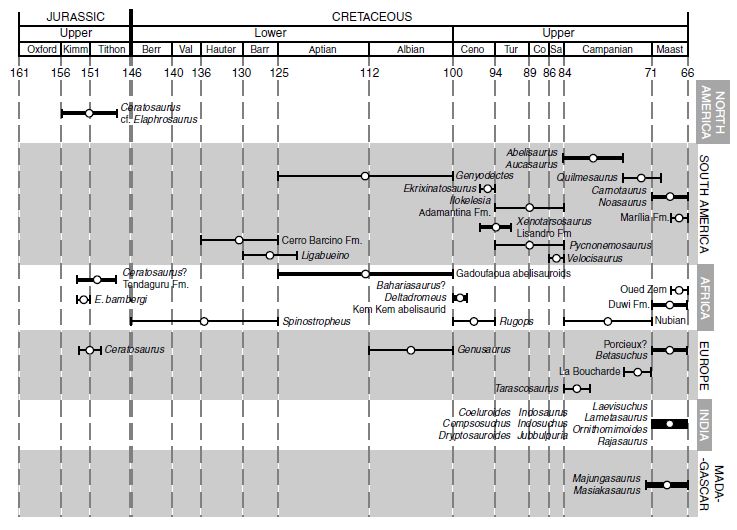
Ceratosaur stratigraphic and geographical distribution, arranged by continent. Error bars reflect uncertainty in age, not time range. The line thickness is based on the number of taxa. No phylogenetic relationships are implied here. Diagram from Carrano & Sampson 2008, timescale from Gradstein et al. (2004). According to Greg Paul (2010), Spinostropheus might actually be Middle Jurassic in age, rather than Early Cretaceous, which would make it the earliest known Ceratosaur Abbreviations: Barr, Barremian; Berr, Berriasian; Ceno, Cenomanian; Co, Coniacian; Fm., Formation; Hauter, Hauterivian; Kimm, Kimmeridgian; Maast, Maastrichtian; Oxford, Oxfordian; Sa, Santonian; Tithon, Tithonian; Tur, Turonian; Val, Valanginian.The following , shows stratigraphic range, illustrating how primitive ceratosaurs conrtinued to co-exuist alongside more advanced forms. |
Ceratosauria phylogeny - a study in phylogenetic controversy
Although the broad brush-strokes of Ceratosaur phylogeny are agreed upon, there is some disagrement when it comes to details of relationships between individual taxa, especially Abelisaurs. This is in part due to the incomplete nature of many species, known only from scrappy remains, and in part due to different cladistic methodologies and approaches. Holtz (1994) was the first to provide a formal cladistic analysis that included both podokesaurs and abelisaurs. He resolved the Ceratosauria into two clades: Coelophysoidea and Abelisauroidea, including Ceratosaurus, Elaphrosaurus and Abelisauridae.
We now know that the Coelophysoidea (i.e., Podokesauridae) as originally defined were a paraphyletic assemblage, but the Abelisauroidea remain valid, and eventually became the redefined Ceratosauria
In any case, there followed a number of other papers. The most notable was Sereno et al 2004 which also described two new species, Spinostropheus gautieri and Rugops primus. These constitute intermediate forms, both morphologically and stratigraphically, between the more basal late Jurassic ceratosaurs Ceratosaurus and Elaphrosaurus, and the more advanced Cretaceous abelisaurs. In this way they arrived at the following cladogram:
The most famous of the abelisaurs, the two-horned Carnotaurus, is here placed as a highly derived form at the summit of a scalloped Abelisauridae, whilst the Noasauridae constitute an unresolved polytomy (i.e. it is not clear which species in the group are related to which). Excluding the poorly resolved Ilokelesia (which seems to jump about almost at random, perhaps due to both its incomplete and its primitive or ancestral nature), the topology (tree shape) is Rugops + (Abelisaurus + (Rajasaurus + (Majungatholus + Carnotaurus))). The last three constitute the Carnotaurinae, the most specialised forms, whereas the eponymous Abelisaurus, which lacks the more dramatic crests and horns characteristic of most other members of the family, belongs with Rugops in a basal position. Indeed, most studiessupport a a close relationship between Carnotaurus and Majungatholus (more properly called Majungasaurus), with Abelisaurus as the outgroup (Sereno 1999; Sampson et al. 2001, Wilson et al 2003; Sereno et al 2004; Tykoski & Rowe 2004; Figs 2, 3).
A radically revised abelisaur phylogeny, based on strict consensus cladogram, was provided by Carrano & Sampson 2008, as follows:
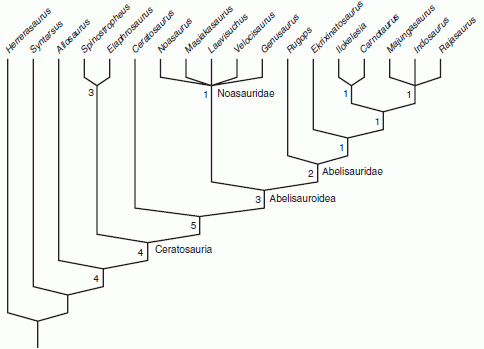
Ceratosaur Cladogram, from Carrano & Sampson 2008 fig 4, the numbers are decay indices. The higher the number, the more robust the cladogram. This doesn't mean that the clades with the highest numbers are more likely to be true, only that they come up most strongly in this particular analysis |
Rather than begining with a single new species, or a few well-known taxa, they incorporated a much larger number of taxa, and this differnt methodology not surprisingly resulte din a different tree topology. Their tree supports a monophyletic Abelisauroidea, Abelisauridae and Noasauridae, but Abelisaurus is no longer a primitive basal form, and the Carnotaurinae dissappear in an unresolved polytomy. Ceratosaurus is intermediate between the three basal African taxa (Elaphrosaurus, Deltadromeus and Spinostropheus) and the Abelisauroidea. There is no internal resolution in the Noasauridae, but within Abelisauridae, Rugops remains the most basal taxon in the latter family. It and Ekrixinatosaurus are successive outgroups to the more derived species (including Abelisaurus, here equal with Carnotaurus and co). Also in contrast to Sereno et al 2004, the three Madagascan and Indian taxa, Majungasaurus, Rajasaurus and Indosaurus, now form a single unnamed clade, supporting the idea of evolution (or more precisely adaptive radiation) through geographic isolation. This makes a lot of sense, as too often cladistics is based on specimens abstracted from geography and stratigraphy, a sif they were idealised platonic forms. Interestingly, early placental mammal evolution similarily appears to have been geographically determined, if molecular phylogeny is anything to go by.
In this particular analysis, Adams consensus revealed that Deltadromeus, Abelisaurus and Aucasaurus acted as 'wildcard' taxa, and so they were pruned to give a more stable cladogram in which Elaphrosaurus, and Spinostropheus are sister taxa (Limusauridae?). The primitive Ilokelesia is also paired with the highly derived Carnotaurus in a rather unlikely clade. The low decay indices (the numbers 1) show how weakly supported some of these results are, although that in itself does not mean they are necessarily incorrect.
Canale et al 2008 return to a more conventional topology (along the lines of Sereno et al 2004), at least as far as abelisaurs go. Their analysis is similarily based on the discovery of a new species, in this case, Skorpiovenator bustingorryi, the most complete abelisaur discovered so far. The Carnotaurines are revived, along with a new clade, Brachyrostra. Rugops remains the most primitive abelisaurid, but Ekrixinatosaurus is now a highly specialised form, paired with Skorpiovenator and Ilokelesia, which is no longer closely related to Carnotaurus. Elaphrosaurus has become a stem-Noasaurid, reflecting the ambiguity among these basal forms.
Finally, a detailed cladistic statistical analysis by Xu et al (2009) using a data matrix incorporating 65 taxa (from Marasuchus to Confusciornis) and 413 morphological characters and the newly described Jurassic ceratosaur Limusaurus inextricabilis, provided the following hypothesis of ceratosaur phylogeny:
The webpage Basal Ceratosauria (in Polish) includes these and many more cladograms, which have been cmbined in the following:
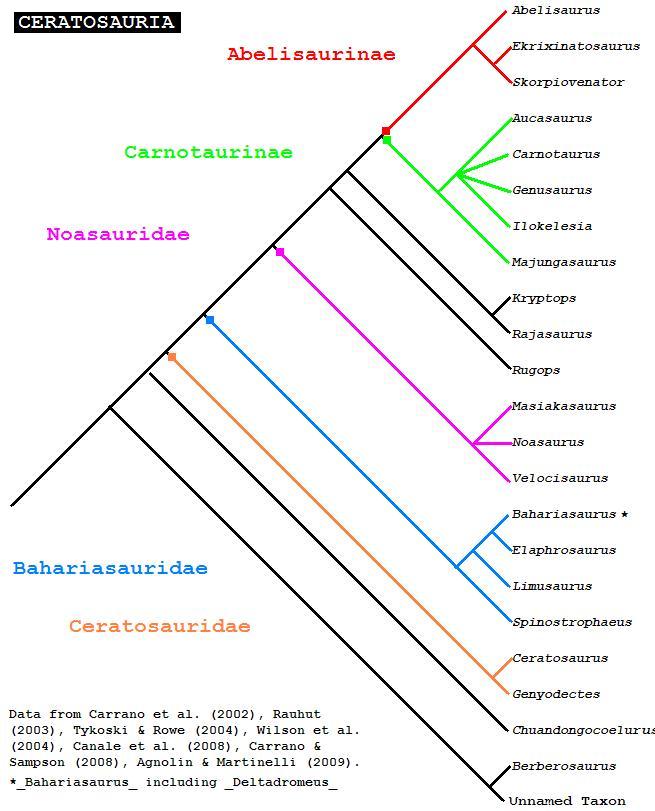
The following Ceratosaur supertree is modified from a dendrogram by Christopher Taylor, 111206, and incorporates the above phylogenies (some incertae sedis taxa removed)
■─
Ceratosauria [Neoceratosauria]
├─
Spinostropheus Xu et al 2009
├─┬─
Elaphrosaurus bambergi Janensch 1920
Xu et al 2009,
Holtz 2000
│ └─
Limusaurus inextricabilis Xu, Clark et al. 2009
Xu et al 2009
└─Ceratosauroidea
WS03
├─
Ceratosaurus nasicornis Marsh 1884
Xu et al 2009,
Holtz 2000
├?─
Deltadromeus agilis Sereno, Dutheil et al. 1996
Xu et al 2009,
Holtz 2000
└─
Abelisauroidea Xu et al 2009
├─
Noasauridae Xu et al 2009
│ ├─
Noasaurus leali Bonaparte & Powell 1980
Xu et al 2009,
N85,
WS03
│ └─
Masiakasaurus knopfleri Sampson, Carrano & Forster 2001
Xu et al 2009,
SCF01
└─
Abelisauridae [Abelisauria]
Xu et al 2009
├─
Rugops Xu et al 2009
└─┬─
Abelisaurus comahuensis Bonaparte & Novas 1985
Holtz 2000 [Abelisaurinae]
WS03
├─┬─
Majungasaurus crenatissimus WS03 (=
Majungatholus atopus Sues & Taquet 1979
WS03,
S03)
│ └─
Rajasaurus narmadensis Wilson, Sereno et al. 2003
WS03 (? = Indosaurus matleyi Huene & Matley 1933
WS03)
└─
Brachyrostra Canale et al 2008 (we have made this a revised Carnotaurinae
WS03)
├─┬─
Ekrixinatosaurus Xu et al 2009
│ └─┬─
Ilokelesia Xu et al 2009
│ └─
Skorpiovenator
└─┬─
Aucasaurus Xu et al 2009
└─
Carnotaurus sastrei Bonaparte 1985
WS03,
Holtz 2000
Descriptions
Ceratosauria Marsh 1884 :
Ceratosaurus, Abelisaurs.
Definition: Ceratosaurus > birds. Padian et al. 1999)
Synonyms: Neoceratosauria
Range: Late Jurassic to Late Cretaceous.
Phylogeny: Averostra : Tetanurae + * : "Limusauridae" + (Ceratosaurus + Abelisauroidea)
Characters: Reduced size of manual phalanges, resulting in relatively weak, or even functionless, hands. This pattern seems to have been reversed in at least a few fingers on the hands of noasaurids)
7 or more sacral vertebrae (Tom Holtz).
Comments: Originally included a number of Triassic and early Jurassic taxa now considered to be basal neotheropoda. The status of the poorly known early Jurassic Sarcosaurus is uncertain, as it has been variously assigned to either the Ceratosauria or the Coelophysoidea, perhaps it is an intermediate form betwene the two. MAK120206
"Limusauridae" (informal taxon name) :
Elaphrosaurus, Limusaurus, ?Spinostropheus
Range: Middle? to Late Jurassic of Afr, Asia, NAm, Eur.
Phylogeny: Ceratosauria : (Ceratosaurus + Abelisauroidea) + * :
Elaphrosaurus + Limusaurus
Comments: It seems that several smaller basal ceratosaurs formed an evolutionary lineage (or several) distinct from the large Ceratosaurids. This seems to be a group of medium-sized, diminutive-armed forms, at least some being toothless omnivores or herbivores. According to Carrano & Sampson 2008, Elaphrosaurus clusters with Spinostropheus; according to Xu et al 2009 with Limusaurus. We might informerly given this new taxon the name "Limusauridae" rather than "Elaphrosauridae" because Limusaurusis known from better material. If Bahariasaurus/Deltadromeus also belongs here (perhaps rather unlikely given the very different forearm proportions and modes of life, although this is not to deny that they might not still be related) then Bahariasauridae von Huene 1948 is an already avaliable name. MAK120217
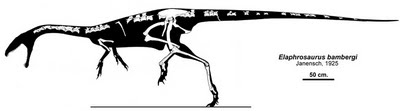
Silhouetted skeletal reconstruction of Elaphrosaurus bambergi by Jaime Headden. |
Elaphrosaurus bambergi Janensch, 1920
Phylogeny: "Limusauridae" : Limusaurus + *
Horizon: Tendaguru beds of Tanzania (Kimmeridgian/Tithonian) ( Late Jurassic of Afr)
Size: length 6.2 meters, weight 210 kg
Comments: Known from a partial skeleton lacking skull, In build it is one of the slenderest and longest (in proprtion to height) of the theropods. Elaphrosaurus was originally considered a typical coelurosaur (using the trem in the the old non-cladistic sense of a wastebasket taxon for any small Jurassic theropod). It was then interpreted as an ancestral Ornithomimid (unlikely in view of the difference in hip structure) and a late surviving coelophysid (Greg Paul). In the absense of a skull, more cannot be determined. The current consensus, confirmed by independent cladistic studies, is that it is a primitive ceratosaur. This has bene further supported with the discovery of a more complete Limusaurus. Isolated remains indicate contemporary French and North American species which may or may not be assigned to this genus MAK010202 120224
Reference: Paul 1988, Xu et al 2009
Limusaurus inextricabilis Xu et al 2009
Horizon: Upper Shishugou Formation in the Junggar Basin of western China. (Oxfordian)
Phylogeny: "Limusauridae" : Elaphrosaurus + *
Comments: the first ceratosaur known from Asia, and the most completely known basal ceratosaur. Cladistic analysis places it as a sister taxon to Elaphrosaurus (Xu et al 2009), so we have united the two in the unofficial family "Limusauridae" (hence the inverted commas). The toothless beak and herbivorous diet make it convergent with Triassic Shuvosaurs, Cretaceous Ornithomimids, and Cenozoic ratites; all being similar in size and general appearance, a remarkable example of convergent evolution among three distinct archosaur lineages. MAK120206
The most extraordinary thing about Limusaurus is what it reveals of the development of the theropod hand and bird wing. To quote Wikipedia:
Limusaurus was a very basal ceratosaur characterized by hands retaining four digits (I-IV), digit I being strongly reduced. It was traditionally thought that the hands of dinosaurs evolved into the wings of birds by the disappearance of the two outward digits (IV and V), in contradiction to embryological studies on birds that showed that the retained digits are the three middle ones (II-III-IV). The hand structure of Limusaurus with its reduced digit I adds more weight to the digit II-III-IV identities for Tetanurae, among which are birds. Previous to the discovery of Limusaurus, theropods were assumed to have progressively evolved reduced digits on the ulnar side of the manus. This concept, known as Lateral Digit Reduction (LDR) is in contrast to Bilateral Digit Reduction (BDR), the reduction on digits on both sides of the hand commonly seen in all other tetrapod groups excluding dinosaurs. However, in Limusaurus, the first digit (Digit I) is strongly reduced, along with other ceratosaurs, suggesting that BDR occurred in their sister group the Tetanurae as well. (Xu et al 2009)
Previously, it was thought that digits I-III were retained in tetanurans as a homology with basal theropods, giving credence to the LDR hypothesis. However, the evidence of BDR in Limusaurus suggests that other non-avian theropods may also have exhibited BDR and the apparent digits I-III in tetanurans may actually be digits II-IV, a previous idea considered by Thulborn and Hamley, but largely ignored in the paleontological community. (Thulborn & Hamley 1982)
Despite the interesting possibilities brought up by Limusaurus, paleontologists are unlikely to stop calling tetanuran digits I, II, and III and switch to calling them II, III, IV. This is because most of their morphological traits resemble those of digits I, II and III of other theropods. It remains possible that bilateral digit reduction occurred in Ceratosauria but not in Tetanurae. The embryology of the bird wing could be explained by a homeotic frameshift of digital identity, as suggested by recent gene expression and experimental data. (Vargas et al 2009)
Links Darren Naish - Limusaurus: awesome and wonderful, with or without the hands
Reference: Xu et al 2009
Images: Theropod Hand Evolution from The Theropod Blog
Spinostropheus gautieri Sereno, Wilson, and Conrad, 2004
Horizon: Tiouraren of Niger - Middle Jurassic of Nth Afr
Phylogeny: Ceratosauria ::: *.
Size: Estimated length: about 4 meters
Comments: Originally Elaphrosaurus gautieri Lapparent, 1960, there seems to be little relation between this species and E. bambergi, apart from the fact that they are both basal ceratosaurs of approximately the same size. Despite the fact that all that is known of this animal is a series of presacral vertebrae, it has been coded in various ceratosaur cladistic analsyses and not surprisingly tends to jump around a lot, although there is agreement on a placement stemward of (more primitive than) the Abelisauroidea. The Tiouraren formation, from which highly distinct dinosaurs such as Jobaria and Afrovenator have been recovered has bene reinterpreted as Middle Jurassic in age (Rauhut & Lopez-Arbarello 2009), although the dating still remains uncertain. MAK120206

Ceratosauridae: ;
Range: Late Jurassic of NAm, Eur, ?Afr
Phylogeny: Ceratosauria : "Limusauridae" + (Abelisauroidea + * : Ceratosaurus nasicornis + Ceratosaurus dentisulcatus)
Comments: Monotypal family of large late Jurassic ceratosaurs, consisting so far only of several species of the single genus Ceratosaurus. These animals existed alongside the better known and more successful Allosaurids. A huge theropod known only from teeth from the Tendaguru of Africa is usually attributed to this group as Ceratosaurus ingens, but could pertain to any large theropod.
Image: Ceratosaurus nasicornis, sketch of skull, from Steele, 1970

Ceratosaurus nasicornis Marsh, 1884
Horizon: Lower and Middle Morrison formation (Kimmeridgian to Early Tithonian) of NAm
Phylogeny: Ceratosauridae : Ceratosaurus dentisulcatus + *.
Size: length about 5.7 meters, weight 500 kg
Comments: For a long time the only well-known ceratosaur, distinguished by its nasal horn/crest. Smaller and less common than it's contemporary Allosaurus. These were ambush predators with large bladed teeth. The deep and heavy tail may have been used for swimming (Paul 2010 p.85). Often (Carrano & Sampson 2008, Xu et al (2009)) considered morphologically intermediate between basal ceratosaurs and abelisaurs. Interesting because it is structurally so much more primitive than its allosaur contemporaries, this medium-sized theropod is characterised by its tall nose horn and smaller preorbital horns in front of the eyes. It also possesses a row of bony nodules down the spine, similiar to those of pseudosuchian thecodonts and the earliest dinosaurs. The skull lightly-constructed skull is armed with quite proportionally large teeth. It is unlikely this animal could bring down large game (in contrast to the Allosaurs); it probably preyed on smaller animals like ornithopods. C. nasicornis is the type species of the genus. C. magnicornis appears to be a synonym (growth stage or intra-specific variation), although it may be a related species or descendant (Paul 1988, Paul 2010) MAK001213 120224
Image: Ceratosaurus nasicornis skeletal reconstruction, redrawn from Greg Paul, from The Real Jurassic Park - Professor Paul Olsen
Ceratosaurus dentisulcatus Madsen and Welles, 2000
Horizon: Upper Morrison formation of Utah (Middle Tithonian)
Phylogeny: Ceratosauridae : Ceratosaurus nasicornis + *.
Size: length 7 meters, weight about 1 tonne
Comments: A large ceratosaur. Quoting Tim Williams (Dinosaur Mailing List): "C. dentisulcatus is the second new Ceratosaurus species named by Madsen and Welles. It appears to be more distinct from C. nasicornis than C. magnicornis is, and so can more reliably be considered a new species. It gets its species name by the distinct grooves (sulci) running down the length of the anterior teeth of the upper jaw. C. dentisulcatus also has fewer maxillary and dentary teeth, and the front of the dentary is strongly curved and upturned. There are also proportional differences in the elements of the skull (such as a lower snout) and postcranium compared to the two other North American Ceratosaurus species. The nasals and lacrimals are not preserved in C. dentisulcatus so its "horns" (assuming it had them) cannot be compared with the other two species." MAK001213 A very similar (Mateus et al 2006), somewhat smaller form is known from Lourinha in Portugal, at the time a large, seasonally dry island with open woodlands, may represent a new species Paul 2010 p.85
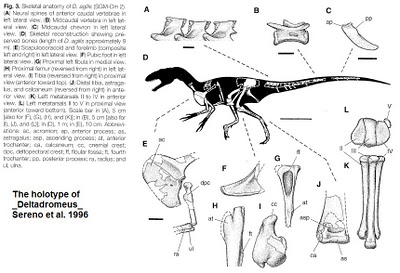
Bahariasaurus ingens Stromer, 1934,
Synonym: or closely related species: Deltadromeus agilis Sereno et al 1996
Range: Mid Cret (Early Cenomanian) of Nth Afr
Phylogeny: Ceratosauria ::: *
Size: 11 to 13.3 meters long, 2.5 to over 4 tonnes in weight (Mortimer 2003, Paul, 2010).
Comments: Bahariasaurus ingens Stromer, 1934, was a gigantic (estimated length 11 meters or more) theropod known from partial postcrania. It was one of several enormous species of dinosaurs unearthed in the Sahara by the German paleontologist Ernst Stromer in the early part of the 20th century; others included Aegyptosaurus, Carcharodontosaurus, and Spinosaurus, as well as the giant crocodilian Stomatosuchus. In 1944, Stromer's entire fossil collection - housed in a Munich museum - was destroyed by an Allied Royal Air Force bombing raid. (Wikipedia). All taht remained were drawings of the spectacular specimens. Forward almost a century and modern paleontologist Paul Sereno andd co-workers undertook their own expeditions to the Sahara, where they found many secimens of what were apparently the same taxa, or at least similar species. For example, the somewhat smaller (8 meters) Deltadromeus agilis Sereno, Dutheil, Iarochene, Larrson, Lyon, Magwene, Sidor, Varricchio, and Wilson, 1996 (certainly author part of a scientific name has grown a lot longer since Stromer's day) appears to be the same animal as Bahariasaurus, probably a juvenile or sub-adult. This was a long-legged, gracile (lightly built) animal, which seems to have been built for speed, but also with unusually large coracoids (part of the shoulder girdle), suggesting long and strong forearms (see illustration of holotype). Originally described as a giant coelurosaur, it was then reinterpreted as a giant Noasaurid (Sereno et al 2004), although most more recent cladistic studies suggest it was actually a basal ceratosaurian (Carrano & Sampson 2008, Carrano et al 2011), with a few dissenters (Cau, et al 2011for example places Deltadromeus in a new clade with -Limusaurus and Masiakasaurus). No skull or complete skeleton is known, and the exact phylogenetic relationship of this animal remains controversial. As to why this part of North Gondwana during the middle Cretaceous featured no less than three quite different gigantic predators is not clear. MAK120210
We have followed Sereno et al 2004 in placing Deltadromeus/Bahariasaurus as a more advanced taxon (above the Ceratosaurid grade), owing to its late appearance. No other basal (primitive) ceratosaur is known from the Cretaceous, and its important not to let ghost lineages multiply out of all proportion. Whether Bahariasaurus is a gigantic noasaur is another matte. it is likely that there were many lines of intermediate-level ceratosaurs in Gondwana at this time. MAK120226
Images: Deltadromeus holotype, from The Theropod Blog
Abelisauroidea Bonaparte 1991 :
Abelisaurus, Carnotaurus, Noasaurus, etc
Phylogenetic definition: The least inclusive clade containing Carnotaurus sastrei Bonaparte 1985 and Noasaurus leali Bonaparte and Powell 1980 (node-based) (Wilson et al 2003) (Paul Sereno - Taxon Search)
Synonyms: Abelisauria
Range: Middle to Late Cretaceous (Aptian/Albian-Maastrichtian) of SAm, India, Madagascar, Europe, Aus
Phylogeny: Ceratosauria ::: Ceratosaurus + (Bahariasaurus + * : Abelisauridae + Noasauridae)
Characters: anterolateral articulation for nasal process, intramandibular socket (Sereno et al 2004), vertebrae with prongs, flange on their femur (Tom Holtz).
Comments: The "crown group" of Createcous ceratosaurs. They are divided into the small noasaurids and the large abelisaurids This recalls the old and outmoded distinction of theropods in general into large (carnosaurs) and small (coelurosaurs), and one may question how accurate such interpretations are. The noasaurs have few if any defining synapomorphies, and may simply represent a primitive assemblage (paraphyletic grade), one particcular lineage of which became the large and specialsed abelisaurids. MAK120217