
Carnotaurus sastrei
artwork courtesy of M. Shiraishi
| Theropoda | ||
| The Vertebrates | Abelisauridae |
| Vertebrates Home | Vertebrate | Vertebrate |
|
Abbreviated Dendrogram
Dinosauria
├─Ornithischia
└─┬─Sauropodomorpha
│
└─Theropoda
├─Herrerasauridae
└─┬─Podokesauridae
└─┬─Dilophosaurus
├─Ceratosauria
│ ├─"Limusauridae"
│ └─┬─Ceratosaurus
│ └─Abelisauroidea
│ ├─Noasauridae
│ └─Abelisauridae
│ ├─Kryptops
│ └─┬─Rugops
│ └─┬─Abelisaurus
│ ├─"Majungasaurinae"
│ │ ├─Majungasaurus
│ │ └─Rajasaurus
│ └─Brachyrostra (= monophyletic Carnotaurinae?)
│ ├─Ekrixinatosaurus
│ └─┬─Ilokelesia
│ └─┬─Skorpiovenator
│ └─Carnotaurini
│ ├─Aucasaurus
│ └─Carnotaurus
└─Tetanurae
└─Avetheropoda
├─Carnosauria
╘═Coelurosauria
└─Aves
|
Contents
Index |
|
Carnotaurus sastrei artwork courtesy of M. Shiraishi |
The Abelisauridae are an important Cretaceous group known mainly from Gondwana. They are also among the more recent dinosaur groups to be discovered, only being unearthed in the 1980s, from rocks of the Latest Cretaceous (Maastrichtian) age in Argentina. In the following decades, other representative oif the group have come to light, whether new discoveries or reinterpretations of older ones, not only in South America but also Madagascar, India, north Africa, and France.
The group includes a number of strange-looking, large to very large, theropods, characterised by a tendency to horns or crests above the eyes (best represented by the iconic horned dinosaur Carnotaurus sastrei), thickening of the skull roof, deep skulls, very lightly built lower jaws, and vestigal, apparently useless arms and hands, reduced so they became even smaller than the tiny forelimbs of the Tyrannosaurs. The abelisaur phylogenetic placement was at one time very questionable. As they combine tyrannosaurid as well as ceratosaur and carcharodontosaur characters, their evolutionary relationship in the past has been difficult to determine. Cladistic analysis however has since unanimously agreed that they are late surviving ceratosaurs. Along with spinosaurs and carcharodontosaurs, abelisaurs make up a unique assemblage of Cretaceous Gondwanan theropods, whose co-existence woudl have been due to niche partitioning. In the later Cretaceous however, abelisaurids were the only large preditors in the southern continents.
MAK990512, MAK120211
 |
Reconstructed Abelisaurus skull, "Dinosaurs of Patagonia" exhibit. Note the rough bone surfaces and projections from the lacrimal and postorbital bones into the eye socket. Photo by , Wikipedia Creative Commons Attribution |
Although skull proportions varied, abelisaurid skulls were generally very tall and often very short. In Carnotaurus, for example, the skull was nearly as tall as it was long. The premaxilla in abelisaurids was very tall, so the front of the snout was blunt, not tapered as seen in many other theropods.
Two skull bones, the lacrimal and postorbital bones, projected into the eye socket from the front and back, nearly dividing it into two compartments. The eye would have been located in the upper compartment, which was tilted slightly outwards in Carnotaurus, perhaps providing some degree of binocular vision. The lacrimal and postorbital also met above the eye socket, to form a ridge or brow above the eye.
Sculpturing is seen on many of the skull bones, in the form of long grooves, pits and protrusions. Like other ceratosaurs, the frontal bones of the skull roof were fused together. Carnotaurines commonly had bony projections from the skull. Carnotaurus had two pronounced horns, projecting outward above the eyes, while its close relative Aucasaurus had smaller projections in the same area. Majungasaurus and Rajasaurus had a single bony horn or dome, projecting upwards from the skull. These projections, like the horns of many modern animals, might have been displayed for species recognition or intimidation (Bonaparte et al 1990, Wilson et al 2003, Tykoski&Rowe 2004)
The forelimbs are known only in the advanced abelisaurids Aucasaurus, Carnotaurus, and Majungasaurus. All had forelimbs which were vestigial. The bones of the forearm (radius and ulna) were extremely short, only 25% of the length of the upper arm (humerus) in Carnotaurus and 33% in Aucasaurus. The entire arm was held straight, and the elbow joint was immobile.Senter 2010
As is typical for ceratosaurs, the carnotaurine hand contained four basic digits. However, it is there that any similarity ends. No wrist bones existed, with the four palm bones (metacarpals) attaching directly to the forearm. There were no finger bones on the first or fourth digits, only one on the second digit and two on the third digit. These two external fingers were extremely short and immobile, and lacked claws Senter 2010
It is unknown if this peculiar forelimb structure applies to more basal abelisaurids, as their forelimbs have not been discovered (Coria et al 2002). More primitive relatives such as Noasaurus and Ceratosaurus had longer, mobile arms with fingers and claws (Agnolin & Chiarelli 2010).
Abelisaurid hindlimbs were more typical of ceratosaurs, with the astragalus and calcaneum (upper ankle bones) fused to each other and to the tibia, forming a tibiotarsus. The tibia was shorter than the femur, giving the hindlimb stocky proportions. There were three functional digits on the foot (the second, third, and fourth), while the first digit, or hallux, did not contact the ground.Tykoski & Rowe 2004. The astragalus and calcaneum (ankle bones) were fused together, and the feet bore three functional digits, with a smaller first digit that did not contact the ground. (Carrano 2007).
Scientists have suggested that the unique skull shape of abelisaurids indicate different predatory habits than other theropods. Whereas most theropods were characterized by long, low skulls of narrow width, abelisaurid skulls were taller and wider, and often shorter in length as well.(Sampson & Witmer 2007) The narrow skulls of other theropods were well-equipped to withstand the vertical stress of a powerful bite, but not as good at withstanding torsion (twisting) (Rayfield et al 2001). In comparison to modern mammalian predators, most theropods may have used a strategy similar in some ways to that of long- and narrow-snouted canids, with the delivery of many bites weakening the prey animal. (Van Valkenburgh & Molnar 2002)
 |
Comparison between the air sacs of Majungasaurus and a recent bird. Graphic by Zina Deretsky, National Science Foundation, via Wikipedia Public Domain |
Scientists have reconstructed the respiratory system of the advanced abelisaurid Majungasaurus based on a superbly preserved series of vertebrae (UA 8678) recovered from the Maevarano Formation. Most of these vertebrae and some of the ribs contained cavities (pneumatic foramina) that may have resulted from the infiltration of avian-style lungs and air sacs. In birds, the neck vertebrae and ribs are hollowed out by the cervical air sac, the upper back vertebrae by the lung, and the lower back and sacral (hip) vertebrae by the abdominal air sac. Similar features in Majungasaurus vertebrae imply the presence of these air sacs. These air sacs may have allowed for a basic form of avian-style 'flow-through ventilation,' where air flow through the lungs is one-way, so that oxygen-rich air inhaled from outside the body is never mixed with exhaled air laden with carbon dioxide. This method of respiration, while complicated, is highly efficient.
The recognition of pneumatic foramina in Majungasaurus, besides providing an understanding of its respiratory biology, also has larger-scale implications for evolutionary biology. The split between the ceratosaur line, which led to Majungasaurus, and the tetanuran line, to which birds belong, occurred very early in the history of theropods. The avian respiratory system, present in both lines, must therefore have evolved before the split, and well before the evolution of birds themselves. This provides further evidence of the dinosaurian origin of birds. (O'Connor & Claessens 2006)
Computed tomography, also known as CT scanning, of a complete Majungasaurus skull (FMNH PR 2100) allowed a rough reconstruction of its brain and inner ear structure. Overall, the brain was very small relative to body size, but otherwise similar to many other non-coelurosaurian theropods, with a very conservative form closer to modern crocodilians than to birds. One difference between Majungasaurus and other theropods was its smaller flocculus, a region of the cerebellum that helps to coordinate movements of the eye with movements of the head. This suggests that Majungasaurus and other abelisaurids like Indosaurus, which also had a small flocculus, did not rely on quick head movements to sight and capture prey.
Inferences about behavior can also be drawn from examination of the inner ear. The semicircular canals within the inner ear aid in balance, and the lateral semicircular canal is usually parallel to the ground when the animal holds its head in an alert posture. When the skull of Majungasaurus is rotated so that its lateral canal is parallel to the ground, the entire skull is nearly horizontal. This contrasts with many other theropods, where the head was more strongly downturned when in the alert position. The lateral canal is also significantly longer in Majungasaurus than in its more basal relative Ceratosaurus, indicating a greater sensitivity to side-to-side motions of the head. (Sampson & Witmer 2007)
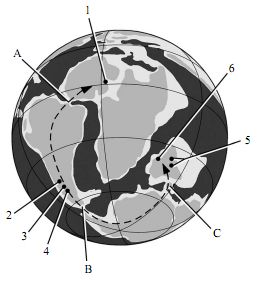 |
Early Late Cretaceous (Early Cenomanian, palaeogeographic map (Scotese 2001) showing all known Late Cretaceous abelisaurid localities (numbers) and key land bridges (letters). The arrow shows the southern high-latitude dispersal route of the 'Africa-first' and 'panGondwana' models of Cretaceous biogeography. The 'Africa-first' model argues that land bridge A was broken in the Early Cretaceous, 30-50 Myr earlier than land bridges B and C; the 'pan-Gondwana' model argues for final separation at land bridge A at the end of the Early Cretaceous. Land bridges: A, Walvis Ridge, Rio Grande Rise; B, Palmer Land Block, South Georgia Island Terrane; C, Kerguelan Plateau, Gunnerus Ridge. Abelisaurid localities: 1, Rugops primus; 2, Aucasaurus garridoi; 3, Abelisaurus comahuensis and Ilokelesia aguadagrandensis; 4, Carnotaurus sastrei; 5, Rajasaurus narmadensis; 6, Majungatholus (= Majungasaurus) atopus. Diagram and caption from Sereno et al 2004). |
The discovery of Abelisaurids in India and Madagascar as well as Argentina inspired theories of paleogeographical migration similar to those postulated for the early evolution of mammals. The most popular is the "Africa-first" model, which supposes that Africa split away from the rest of Gondwana first, the remainder of the supercontinent being unified by two land bridges until the Late Cretaceous (figure 1, land bridges B, C). This is contradicted both by palaeogeographic reconstructions (which have several landmasses separated by intervening seaways early in the Cretaceous (Smith et al. 1994; Roeser et al. 1996), as well as paleontological evidence such as notosuchian crocodylomorphs that show a close connection between of mid-Cretaceous African and South American crocodylomorphs, carcharodontosaur dinosaurs, and so on. An alternative model is the "pan-Gondwana" hypothesis, where three narrow and probably intermittent, passages connected major Gondwanan landmasses during the Early Cretaceous (figure 1, land bridges A-C). These were severed during a relatively brief interval during the middle Cretaceous. This differs from the "Africa-first" model in the presence of a trans-Atlantic passage (land bridge A). Similarity of faunas across continets during this time (including Abelisaurs such as Rugops even in continental Africa) supports a narrow time interval for their permanent separation ( Sereno et al 2004)
Significantly however, recent research (Carrano & Sampson 2008) supports the existence of an endemic central Gondwanan (India and Madagascar) clade distinct from the West Godwnana (South American) forms. It is likely that this group evolved in the late Cretaceous following separation of the continents, although the West Gondwanan clade (Brachyrostra) is known even from the middle Cretaceous
Abelisauridae Bonaparte and Novas 1985
Abelisaurus, Carnotarus
The most inclusive clade containing Carnotaurus sastrei Bonaparte 1985 but not Noasaurus leali Bonaparte and Powell 1980, Coelophysis bauri (Cope 1889), or Passer domesticus (Linnaeus 1758). (stem-based) (Paul Sereno - Taxon Search)
Middle to Late Cretaceous (Aptian/Albian-Maastrichtian) of SAm, India, Madagascar, Europe
Abelisauroidea : Noasauridae + * : Rugops + (Abelisaurus + Carnotaurinae)
Very rugose (horny) texture of the bones of the face: this would have supported some sort of softer tissue, but whether keratin (horn-like structures) or some thickened skin is not ceratin; Short rounded snouts; Thickened skull roof; Squat thickened teeth; Forearms highly reduced: the ulnae and radii are practically no more than carpals; Relatively short and stocky hindlimbs. (Tom Holtz), groove-and-pit ornamentation, basic abelisaurid skull design (Sereno et al 2004).
A remarkably succesful clade of large theropods, known from South America, India, Madagascar, and Northern Africa. No doubt future discoveries will reveal and even greater distribution. During the Middle Cretaceous they not only co-existed with giant charcharodontosaur and spinosaur theropods, but actually outlived them, becoming the apex predators of every continet except Asia-America. These were large preditors that filled the same ecological role in middle and late Cretaceous Gondwana as Allosaurs and Tyrannosaurs did during the Cretaceous of the northern supercontinent (Laurasia). An original division of abelisaurids into abelisaurines, with rather long, flat skulls that resemble in some ways the skulls of tyrannosaurids, and carnotaurines, with short, deep, ornamented skulls, has not been confoirmed by cladistic analysis and so is rejected here. It seems instead that the development of horn-like structures and differential cranial thickening appear to be convergently acquired within Abelisauridae. These short thin crests or horns are very different to the dilophosaur crests. All abelisaurs appear to have had side-to-side expanded "cheeks," like those of tyrannosaurids. Whilst Abelisaurus and Carnotaurus are the two best known genera, there are a number of other forms that have recently been placed here, and more are being discovered all the time. Abelisaurids had the greatest reduction in forelimbs among the theropods, even tyrannosaurs had proprtionally larger arms. The vestigal arms (reminiscent of the tiny hind limbs of early whales) would have been useless in grappling or any other purpose. As with tyrannosaurs, the powerful skulls and jaws were used to subdue prey.
MAK990512 MAK120210
 |
Kryptops palaios, known fossil elements, after Sereno & Brusatte 2008. Artwork by Nobu Tamura, Wikipedia, GNU Free Documentation/Creative Commons Attribution |
Gadoufaoua, on the western edge of the Ténéré Desert
Elrhaz Formation
Aptian–Albian
Abelisauridae : (Rugops + (Abelisaurus + "Majungasaurinae" + Brachyrostra/Carnotaurinae)) + *
secondary wall on the maxilla that obscures the antorbital fenestra in lateral view, articular trough for the nasal on the maxilla is narrower and less developed, which is a primitive condition. Texturing of the external surface of the maxilla composed of shorter grooves than on other abelisaurids. The sacrum and ilium are also more primitive than in Majungasaurus and Carnotaurus, sacrum with only five vertebrae, and ilium with a relatively deeper preacetabular process. (Sereno & Brusatte 2008 p.18)
Known from a maxilla (part of upper jaw) and postcranial elements, this is the oldest known and most primitive Abelisaurid. Features of the maxilla such as vascular grooves and a narrow antorbital fossa place Kryptops among the abelisaurids (Sereno & Brusatte 2008). This is one of those instances, useful in vertebrate paleontology when dealing with incomplete specimens, where a sintgle bone can identify a species. As many distinct lineages of abelisaurids co-existing in this area is perhaps not likely, it is possible that Kryptops ("hidden face") was the ancestor, or at least a very close cousin, of Rugops ("rugged face"). Both lived in the same geographic region, and the latter only ten million or so years later. This animal was a contemporary of theprimitive carcharodontosaur Eocarcharia dinops, both abelisaurids and carcharodontosaurs appeared at the same time and would evolve in parallel in Gondwana during the middle Cretaceous
MAK120226
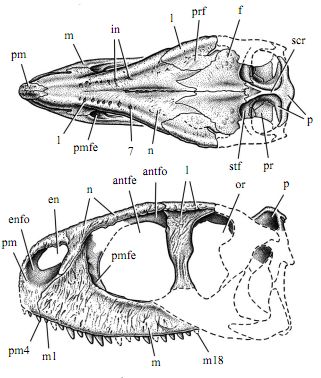 |
Rugops primus. skull length 31.5 cm Already in this basal form the characteristic short high morphology has developed, but not yet the thickening of the skull roof and the ridges and crests found in some highly derived species. 1-18, tooth positions or number of nasal invagination; antfe, antorbital fenestra; antfo, antorbital fossa; en, external naris; enfo, external narial fossa; f, frontal; fo, foramen; in, invagination; l, lacrimal; m, maxilla; n, nasal; or, orbit; p, parietal; pm, premaxilla; pmfe, promaxillary fenestra; pr, prootic; prf, prefrontal; scr, sagittal crest; se, septum; stf, supratemporal fossa Drawing from Sereno et al 2004 fig. 3. |
In Abangharit, Niger Republic
Echkar Formation
Cenomanian
Abelisauridae : Kryptops + (Abelisaurus + "Majungasaurinae" + Brachyrostra/Carnotaurinae) + *)
small fenestra (opening) in the skull roof between the prefrontal, frontal, post-orbital and lacrimal, and a row of seven small invaginated depressions on the dorsal surface of each nasal. Sereno et al 2004
Known from a partial skull lacking the palate and most of the bones along the side of the face behind the eyes, this is one of the earliest abelisaurids with textured external skull surfaces. It consistently resolves as a basal form in cladistic analyses
Unnamed Node: Higher Abelisaurids
Cenomanian-Maastrichtian of SAm, India, Madagascar
Abelisauridae ::: (Abelisaurus + "Majungasaurinae" + Brachyrostra/Carnotaurinae) + *
thickened skull roof Sereno et al 2004
All analyses agree that Abelisaurus and Carnotaurines share a common ancestor above the level of Rugops, regardles sof whether or not Carnotaurinae are a natural group. The Cenomanian appears to have been an important time in abelisaurid evolution.
MAK120210
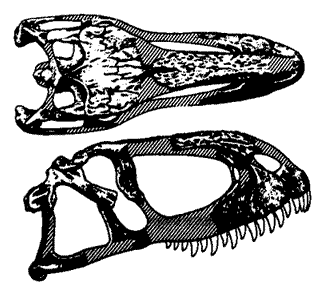 |
Abelisaurus comahuensis, type specimen, MC 11098. Drawing by Greg Paul, Predatory Dinosaurs of the World, p.284 |
Abelisaurus comahuensis Bonaparte and Novas, 1985
a monospecific Abelisaurinae (Paul 1988), Genyodectes?
Allen Formation
Rio Negro, Argentina
early or mid Campanian
Phylogeny: Abelisauridae : Kryptops + (Rugops + ("Majungasaurinae" + Brachyrostra/Carnotaurinae + *))
Known only from a large, nearly complete skull, Abelisaurus was unusual when first discovered in view of its late occurance and primitive megalosaur-like features. The eye socket's orbital bar and simple lower cheek bones indicate it is closely related to Carnotaurus. Genyodectes serus Woodward, 1901, also from the latest Cretaceous of Argentina, and based on part of the front end of some theropod jaws, may be closely related, if not synonymous.
MAK990512
The basal position of Abelisaurus assumes that the rugged horned and crested form only evolved once (Carnotaurinae) (Sereno et al 2004, see also cladogram in Wilson et al 2003). Current cladistic analaysis indicates there was a lot of parallelism in abelisaurid evolution (Carrano & Sampson 2008, Canale et al 2008), since both horned and hornless forms evolved among higher taxa, there is no reason why Abelisaurus should be discriminated against. Therefore we have moved this taxon to the crown group Abelisaurids
MAK120211
Indosaurus, Majungasaurus, Rajasaurus
 |
Majungasaurus crenatissimus, life reconstruction .Artwork by Nobu Tamura, Wikipedia, GNU Free Documentation/Creative Commons Attribution |
Maastrichtian of India & Madagascar
Abelisauridae : Kryptops + (Rugops + (Abelisaurus + Brachyrostra/Carnotaurinae + * : Majungasaurus + Rajasaurus))
analysis by Carrano & Sampson 2008 recovered a clade of large, powerfully-built abelisaurids, that populated Central Gondwana (India and Madagascar ). As it doesn't seem to have a name we here informally refer to this lineage as Majungasaurinae, after the best known species, Majungasaurus crenatissimus
The Majungasaurines represent a distinct lineage to the short-headed, highly derived West Gondwanan forms. The total tibia-femur length is notably short compared to other similarly sized theropods, indicating a relatively slow moving animal. They also lack the unique caudal morphology and powerful leg muscles of the advanced West Gondwanan forms (Persons & Currie 2011 pp.1-2). These differences may reflect different faunal compositions. For example, ornithopods are unknown from Central Gondwana. These powerfully built abelisaurs therefore would have preyed on large and slow titanosaurs (sauropods) rather than smaller and faster ornithopods, All members of this unnamed clade are currently known only from the Maastrichtian, implying a ghost lineage of thirty or forty million years.
Majungasaurus crenatissimus (Depéret, 1896)
Megalosaurus crenatissimus Depéret, 1896, Dryptosaurus crenatissimus (Depéret, 1896) , Majungatholus atopus Sues and Taquet, 1979
 |
|
Majungasaurus crenatissimus, Field Museum, Chicago. Photo by Ed Bierman, Wikipedia Creative Commons Attribution |
Maevarano Formation
Mahajanga Basin, northwestern Madagascar
Maastrichtian
"Majungasaurinae" : Rajasaurus + *
thickened nasals, a horn on the frontal, a parietal prominence, and pneumatic chambers.
average 6-7 meters in length (Krause et al 2007), reached lengths of more than 8 meters
average weight 750 (Paul 2010) to 1100 kilograms, although large specimens heavier ((Sampson & Witmer 2007)
This theropod was for a long time considered to be a large pachycephalosaurid, called Majungatholus atopus. It is distinguished by thickened nasal bones, a horn on the frontal bone, and a parietal (skull bone near the rear) prominence. The earlier named Majungasaurus crenatissimus, originally based on an incomplete dentary from the same time and place, has a curved mandible is very like that of Carnotaurus. Recent excavations undertaken by the Mahajanga Basin Project resulted in abundant and spectacular remains of this theropod, including multiple specimens of various size and presumably age ranges, and covering the great majority of the skull and skeleton, make Majungasaurus among the best known of Gondwanan theropods, and conclusively placing it within Abelisauridae . The confused taxonomy is also resolved, with Majungasaurus crenatissimus as the valid name for this theropod (Krause et al 2007). Previously considered closely related to Carnotaurus (Wilson et al 2003), new analysis places it separate from the South American forms (Carrano & Sampson 2008, Canale et al 2008)
Matching tooth marks on bones show that this species sometimes fed on its own kind (Rogers et al 2007) (Cannibalistic behaviour had previously been attributed to Coelophysis bauri, but this is now known not to have been the case)
The following material is copied or adapted from the Wikipedia page on Majungasaurus
 |
The face of Majungasaurus crenatissimus Artwork by Deviant Paleoart, Wikipedia Creative Commons Attribution |
The skull of Majungasaurus is exceptionally well-known compared to most theropods. Like other abelisaurid skulls, its length was proportionally short for its height, although not as short as in Carnotaurus. The skulls of large individuals measured 60-70 centimeters (24-28 in) long. The tall premaxilla (frontmost upper jaw bone), which made the tip of the snout very blunt, was also typical of the family. The nasal bones were extremely thick and fused together, with a low central ridge running along the half of the bone closest to the nostrils. A distinctive dome-like horn protruded from the fused frontal bones on top of the skull as well. In life, these structures would have been covered with some sort of integument, possibly made of keratin. Computed tomography (CT scanning) of the skull shows that both the nasal structure and the frontal horn contained hollow sinus cavities, perhaps to reduce weight (Sampson & Witmer 2007). The hollow cavity inside the frontal horn of Majungasaurus would have weakened the structure and probably precluded its use in direct physical combat, although the horn may have served a display purpose (Tykoski & Rowe 2004). While there is variation in the ornamentation of Majungasaurus individuals, there is no evidence for sexual dimorphism.(Sampson & Witmer 2007). The teeth were typical of abelisaurids in having short crowns, although Majungasaurus bore 17 teeth in both the maxilla of the upper jaw and the dentary of the lower jaw, more than in any other abelisaurid except Rugops (Smith2007).
Abelisaurids may instead have been adapted for a feeding strategy more similar to modern felids, with short and broad snouts, that bite once and hold on until the prey is subdued. Majungasaurus had an even broader snout than other abelisaurids, and other aspects of its anatomy may also support the bite-and-hold hypothesis. The neck was strengthened, with robust vertebrae, interlocking ribs and ossified tendons, as well as reinforced muscle attachment sites on the vertebrae and the back of the skull. These muscles would have been able to hold the head steady despite the struggles of its prey. Abelisaurid skulls were also strengthened in many areas by bone mineralized out of the skin, creating the characteristic rough texture of the bones. This is particularly true of Majungasaurus, where the nasal bones were fused and thickened for strength. On the other hand, the lower jaw of Majungasaurus sported a large fenestra (opening) on each side, as seen in other ceratosaurs, as well as synovial joints between certain bones that allowed a high degree of flexibility in the lower jaw, although not to the extent seen in snakes. This may have been an adaptation to prevent the fracture of the lower jaw when holding onto a struggling prey animal. The front teeth of the upper jaw were more robust than the rest, to provide an anchor point for the bite, while the low crown height of Majungasaurus teeth prevented them from breaking off during a struggle. Finally, unlike the teeth of Allosaurus and most other theropods, which were curved on both the front and back, abelisaurids like Majungasaurus had teeth curved on the front edge but straighter on the back (cutting) edge. This structure may have served to prevent slicing, and instead holding the teeth in place when biting.(Sampson & Witmer 2007)
Majungasaurus was the largest predator in its environment, while the only known large herbivores at the time were sauropods like Rapetosaurus. Scientists have suggested that Majungasaurus, and perhaps other abelisaurids, specialized on hunting sauropods. Adaptations to strengthen the head and neck for a bite-and-hold type of attack might have been very useful against sauropods, which would have been tremendously powerful animals. This hypothesis may also be supported by the hindlegs of Majungasaurus, which were short and stocky, as opposed to the longer and more slender legs of most other theropods. While Majungasaurus would not have moved as fast as other similar-sized theropods, it would have had no trouble keeping up with slow-moving sauropods. The robust hindlimb bones suggest very powerful legs, and their shorter length would have lowered the animal's center of gravity. The tibia (lower leg bone) of Majungasaurus was even stockier than that of its relative, the sprinter Carnotaurus, with a prominent crest on the knee (Carrano 2007). Thus Majungasaurus may have sacrificed speed for power (Sampson & Witmer 2007) Majungasaurus tooth marks on Rapetosaurus bones confirm that it at least fed on these sauropods, whether or not it actually killed them. (Rogers et al 2007)
Rajasaurus narmadensis Wilson, Sereno, Srivastava, Bhatt, Khosla, and Sahni, 2003
Lametasaurus, Indosaurus?
Narmada Valley, Rajasthan, India
Infratrappean Beds of the Lameta Formation
Maastrichtian
single nasal-frontal horn, the elongated proportions of its supratemporal fenestrae (holes in the upper rear of the skull), and the form of the ilia (principle bones of the hip) which feature a transverse ridge separating the brevis shelf from the hip joint. (Wilson et al 2003)
7.6-9 m long, 2.4 m high (Wikipedia)
about 3 to 4 tons (Wikipedia)
the first Indian theropod preserving associated cranial and postcranial remains; known from an almost complete skull and partial skeleton . The skull was short, measuring 60 cm (23.6 in) in length, and bore a distinctive low rounded horn or crest, made of outgrowths from the nasal and frontal bones. The configuration of its nasal bones and possession of a growth ("excrescence") on its frontal bone places it among the carnotaurine abelisaurids (Wikipedia), although the presence of these features developed independently among the abelisauruids (hence Abelisaurus itslef may not be as basal as usually portrayed (Carrano & Sampson 2008)). Rajasaurus seems to have been a fairly heavily built animal. Indosaurus matleyi from the Lower Lameta Group, Madhya Pradesh, India (mid-late Maastrichtian ) may be a synonym or at least a very closely related species (which would mean renaming Rajasaurus), although the type material is lost. It was known from a skull roof and other cranial and post-cranial elements, with frontal bones massively thickened into what appear to be the bases of horn cores above the eyes. In any case, proper identification of these Indian forms is hampered by poor preservation Carrano & Sampson 2008. The contemporary Indosuchus, known from very scrappy remains, was a smaller, longer-legged animal (Paul 2010).
This was a very large animal, although Greg Paul's (2010) length estimate of 11 meters would seem to be excessive
Presumably the weight estimate is for the larger end of the size range.
Carnotaurus, Skorpiovenator
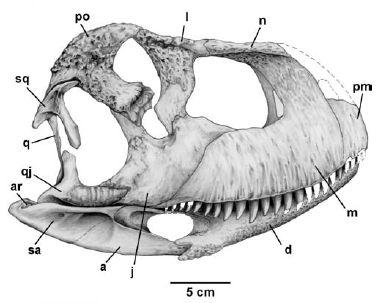 |
Skull of Skorpiovenator bustingorryi, showing the short high shape typical of advanced abelisaurids. Compare with the more familiar, but probably distorted, shape of Carnotaurus, below. d Dentary, a angular, sa surangular, ar articular, pm premaxilla, m maxilla, n nasal, j jugal, l lachrymal, po postorbital, qj quadratojugal, q quadrate, sq squamosal. From Canale et al 2008 fig.2. |
Carnotaurinae: The most inclusive clade containing Carnotaurus sastrei Bonaparte 1985 but not Abelisaurus comahuensis Bonaparte and Novas 1985. (stem-based) (Paul Sereno - Taxon Search).
However Carrano & Sampson 2008's findings collapse traditional abelisaurid phylogeny and present the Carnotaurinae as paraphyletic; the relationship between Carnotaurus, Abelisaurus, and other higher taxa dissapears in an unresolved polytomy. A sim8ilar but not synonymous stem-based clade would be Brachyrostra: all abelisaurids more closely related to Carnotaurus sastrei than to Majungasaurus crenatissimus (Canale et al 2008). For now, we have chosen to present Abelisaurus, Carnotaurus, and Majungasaurus as representing three equally distinct clades.The position of Ekrixinatosaurus is uncertain; it may represent a more basal form than all three, or a stem member of the Carnotaurus clade (Brachyrostra/Carnotaurinae)
Late Cretaceous ( Late Cenomanian/Early Turonian-Maastrichtian) of SAm
Abelisauridae : Kryptops + Rugops + (Abelisaurus + "Majungasaurinae" + * : Ekrixinatosaurus + (Ilokelesia + (Skorpiovenator + Carnotaurini)))
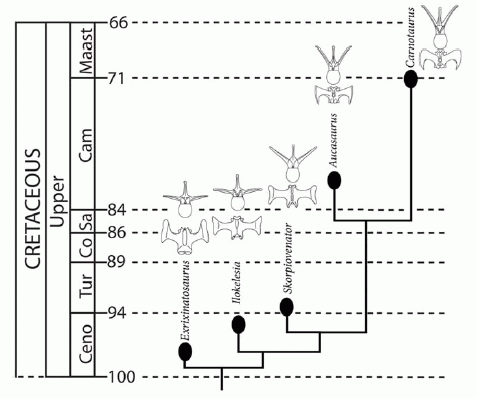
The Brachyrostra ("short snouts") are a postulated clade of abelisaurs geographically limited to South America, showing that abelisaurids divided early on along geographical lines, which fits with the timing of the break up of Gondwana. Distinct from both the Abelisaurines and the Majungasaurines, these advanced abelisaurs were characterised by a shortening of the skull and hyperossification (thickening of the bone) of the skull roof seem to be correlated with a progressive enclosure of the orbit. This may be related to shock-absorbing, implying head-butting intra-specific behaviour. Canale et al 2008. Phylogenetic analysis of Canale et al 2008, recovered three species ina stratigraphic sequence, the newly discovered Skorpiovenator, the Carnotaurus-like Ekrixinatosaurus, and the poorly known Ilokelesia as part of a newly discovered clade that is the sister group to the Carnotaurini (Aucasaurus + Carnotaurus). This contradicts Sereno et al 2004, for whom Ilokelesia is a basal form, and Carrano & Sampson 2008, who place Ekrixinatosaurus intermediate between Rugops and higher Abelisaurs, although the latter authors did recover a connection between Ilokelesia and Carnotaurus. Persons & Currie 2011 in their study of abelisaurid tail morphology note that these five taxa form both a morphological and a stratigraphic series, especially in view of their close geographic proximity, and we have followed this hypothesis here. Although Canale et al 2008 find that in Skorpiovenator and Ekrixinatosaurus horns consist of dorsally inflated postorbital bones, distinct from the frontal horns of Carnotaurus, such reversals would not be unusual in any evolutionary (anagenetic) sequence. Advanced members of this clade became progressively more cursorial, culminating in Carnotaurus, the fastest known large theropod (Persons & Currie 2011)
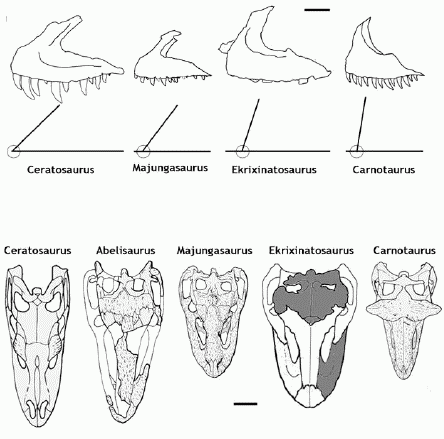 |
Determining phylogentic placement from oinly a few partial elements. The maxilla of Ekrixinatosaurus possesses a dorsally projected ascending ramus and a short rostral ramus, suggesting a relatively high skull, most similar to that of Skorpiovenator and Carnotaurus rather than the longer-headed Majungasaurus or Rugops. The relationship between the length of the maxilla and the width of the skull roof also implies a short skull, intermediate in length between those of Majungasaurus and Carnotaurus, and similar to Skorpiovenator. All these taxa belong to the Brachyrostra or short-headed clade. Diagram and some of text from Juarez Valieri et al 2011 |
Upper Candeleros Formation
Patagonia, Argentina
Early Cenomanian
Brachyrostra : (Ilokelesia + (Skorpiovenator + Carnotaurini)) + *
Originally described as 7 - 8 meters in total length (Calvo et al. 2004), revised estimates put this at 10 to 11 meters, making this the largest known abelisaur (Juarez Valieri et al 2011)
Ekrixinatosaurus (the name means "explosion-born lizard," as it was discovered during a blasting operation (ref Thescelosaurus)) is based on a partial skeleton and skull elements. It shares a mixture of characters with several other abelisaurids, highlighting the homoplastic, probably ontogenetic, nature of many of these traits (Carrano & Sampson 2008). Either a fairly large abelisaur with an oversized head, or the postcrania from one indivdual along with skull fragments of a much lartger one. A particularly robust animal, a contemporary of the even larger carcharodontosaurid Giganotosaurus. As a result of the post-Cenomanian Carcharodontosaurs, both animals being the largest known representatives of their respective clades. (Juarez Valieri et al 2011). Stratigraphically this is earliest of the Brachyrostra, and as its stocky build and lacks of advanced cursorial adaptations means it was not a fast moving animal (Persons & Currie 2011 p.2). Variously considered a derived (Canale et al 2008) or a primitive (Carrano & Sampson 2008, Persons & Currie 2011) species, the combination of brachyrostran head and "Majungasaurine" body and locomotary traits, along with early stratigraphic position (the earliest well known abelisaurid) support its more basal position.
Ilokelesia aguadagrandensis Coria and Salgado, 2000
Río Limay Formation, Neuquén Group
Neuquén Province, Argentina
Late Cenomanian
Brachyrostra : Ekrixinatosaurus + ((Skorpiovenator + Carnotaurini) + *)
Known from fragmentary material, and initially thought to be a basal abelisauroid due to the retention of basal characteristics, it now seems to a more advanced animal, representing an early stage in the brachyrostral lineage Canale et al 2008 and/or related to Carnotaurus (Carrano & Sampson 2008) or may be an intermediate form between Ekrixinatosaurus and Skorpiovenator (Persons & Currie 2011) The base of the tail is exceptionally broad (Paul 2010)
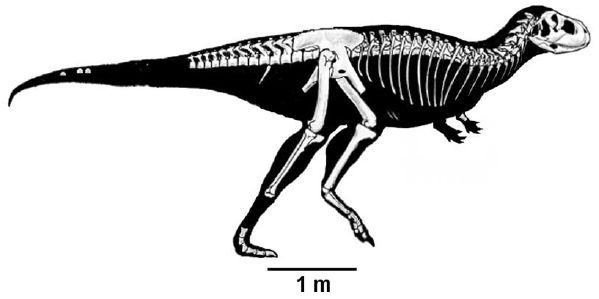 |
Skorpiovenator bustingorryi, reconstruction skeleton, from Canale et al 2008 fig.1. |
lower Huincul Formation
Patagonia
Late Cenomanian/Early Turonian)
Brachyrostra : Ekrixinatosaurus + (Ilokelesia + ( Carnotaurini + *))
Known from a complete skull and most of the skeleton, an dnamed after the large number of scorpions at the fossil locality, this long legged form represents the beginning of the tendency towards advanced cursorial ability rapid sprinting and - by implication, as these were the largest predators of their environment - pursuit that reached its culmination in the avanced carnotaurini (Persons & Currie 2011)
Based on similarities between Skorpiovenator and carcharodontosaurid tooth morphology, it is quite likely that stratigraphically younger isolated teeth originally referred as Carcharodontosauridae belong to abelisaurids Canale et al 2008. Since no post-Cenomanian Carcharodontosaurs are otherwise known, this implies that that group became extinct at the end of the Cenomanian, their ecological role being replaced by large abelisaurs, although their smaller (average 6 meters) cousins or descendents the neovenatorids continued to flourish in the medium megapredator role.
 |
Carnotaurus sastrei, cast of skeleton, Canadian Museum of Nature, Ottawa. Photo by D. Gordon E. Robertson, Wikipedia GNU Free Documentation/Creative Commons Attribution |
Aucasaurus, Carnotaurus
Late Cretaceous (Campanian-Maastrichtian) of SAm
Brachyrostra : Ekrixinatosaurus + (Ilokelesia + (Skorpiovenator + * : Aucasaurus + Carnotaurus))
Canale et al 2008 place Carnotaurus and Aucasaurus as sister taxa in the tribe Carnotaurini. This was the only West Gondwanan abelisaurid lineage to survive to the end of the Cretaceous. In comparison to other Abelisaurids, the legs are long and gracile. This lineage of large theropods specialise din fast pursuit, unlike the more heavily built central Gondwanan clade. As Persons & Currie 2011 explain in the abstract:
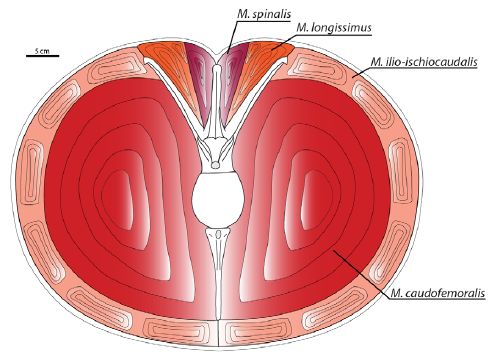
In the South American abelisaurids Carnotaurus sastrei, Aucasaurus garridoi, and, to a lesser extent Skorpiovenator bustingorryi, the anterior caudal ribs project at a high dorsolateral inclination and have interlocking lateral tips. This unique morphology facilitated the expansion of the caudal hypaxial musculature at the expense of the epaxial musculature. Distinct ridges on the ventrolateral surfaces of the caudal ribs of Aucasaurus garridoi are interpreted as attachment scars from the intra caudofemoralis/ilio-ischiocaudalis septa, and confirm that the M. caudofemoralis of advanced South American abelisaurids originated from a portion of the caudal ribs Digital muscle models indicate that, relative to its overall body size, Carnotaurus sastrei had a substantially larger M. caudofemoralis than any other theropod yet studied. In most non-avian theropods, as in many extant sauropsids, the M. caudofemoralis served as the primary femoral retractor muscle during the locomotive power stroke. This large investment in the M. caudofemoralis suggests that Carnotaurus sastrei had the potential for great cursorial abilities, particularly short-burst sprinting. However, the tightly interlocking morphology of the anterior caudal vertebrae implies a reduced ability to make tight turns.
 |
 |
Chronostratigraphy and hypothesized phylogeny of South American Abelisauridae with representative caudal vertebrae for each in anterior and dorsal views. Note: although each taxon is demarked by a separate branching event, given the close geographic and temporal proximities of these taxa, combined with the unlikelihood that multiple other as-yet-unknown large-bodied carnivorous abelisaurids were coexistent, it is probable that some of these taxa have a direct anagenetic relationship with others. Caption and figure from Persons & Currie 2011 fig.6 Creative Commons Attribution. |
Cross-section through the tail of Carnotaurus sastrei showing caudal vertebra 6 and accompanying musculature. Note: the cross-section is an anatomical abstraction and depicts the neural arch and chevron in the same vertical plane. Caption and figure from Persons & Currie 2011 fig.5. |
 |
Aucasaurus garridoi - Mounted skeleton reconstruction Photo by Kabacchi, Wikipedia, Creative Commons Attribution |
Aucasaurus garridoi Coria, Chiappe, and Dingus, 2002
Anacleto Formation
Argentina
Early Campanian
Carnotaurini : Carnotaurus + *
Related to but smaller than Carnotaurus, and lacking the head crests, with a longer, lower skull with bumps instead of horns. The arms were also somewhat longer. Based on an almost complete skeleton, including soft tissue impressions about the hips. (Ref - Thescelosaurus). Despite the complete fossil, this animal remains poorly known.
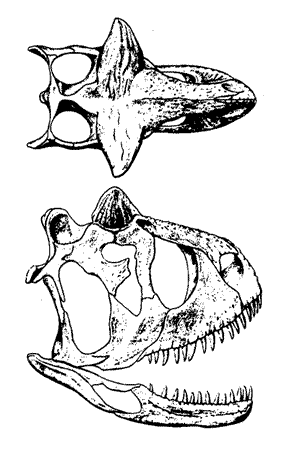 |
Carnotaurus sastrei, type specimen, CH894. The exceptionally short proportions of the skull, even by abelisaur standards, may be the result of post-moortem deformation. Drawing by Greg Paul, Predatory Dinosaurs of the World, p.284. |
Gorro Frigio Formation
Chubut, Argentina
Maastrichtian
Carnotaurini : Aucasaurus + *
length 7.5 to 9 meters (Mazzetta et al 1998, Paul 2010)
weight 1500 to 2000 kilograms .(Mazzetta et al 1998, Paul 2010)
Known remains consist of a skull and and almost complete skeleton with skin impressions.
This theropod is known from a virtually complete skeleton (lacking only the hind feet and end of the tail), a skull with extensive skin impressions. The skull is very high and short with two stout horn-cores, which in life were the basis of extended horns. this has inspired the strange name "flesh-eating bull". The horns were probably quite blunt and used for intraspecific combat (head butting etc). The eyes are quite small (much of the orbit cut off by a bar of bone) although Greg Paul raises the possibility the deep snout may have contained nasal organs. Perhaps unlike most other theropods this dinosaur hunted by smell rather than sight. The skin empressions show this animal to have had widely-spaced rows of nonbony scutes. The hands and arms are so small as to be practically useless.
Carnotaurus seems to have had a highly kinetic skull and could gape widely to swallow large objects. A prokinetic hinge rostral to the orbits would have allowed (as in birds) elevation of the rostrum while the eyes could remain fixed on prey. The vaulted skull gave a fast, rather than a strong, bite. Carnotaurus was envisioned as an active predator of fast, small prey.. These large speialised animals were among the last of the South American abelisaurids
It is possible the distinctive shape of the type skull of Carnotaurus is due to distortion. Mark Goodwin et al., in their initial presentation on Majungatholus (=Majungasaurus), suggested that the deep and narrow skull shape of Carnotaurus owed itself to compaction and might not have been natural. (SVPCA - Darren Naish)
This was the fastest large theropod, with the largest M. caudofemoralis in proportion to its size (Persons & Currie 2011). It represented the culmination of the West Gondwanan brachyrostran lineage.
checked ATW030913