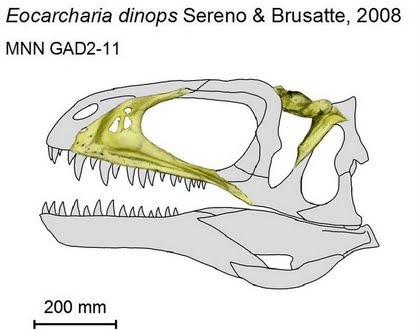
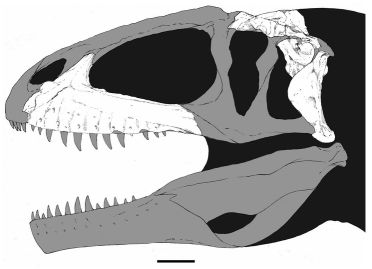
Flesh reconstruction and line drawing of the skull of Acrocanthosaurus atokensis (NCSM 14345) in left lateral view. Hatched lines represent missing bone. A, angular; aof, antorbital fenestra; AR, articular; D, dentary; emf, external mandibular fenestra; iop, intraorbital process of postorbital; J, jugal; L, lacrimal; lpr, lacrimal pneumatic recess; ltf, lateral temporal fenestra; M, maxilla; mf, maxillary fenestra; N, nasal; o, orbit; ob, orbital boss of postorbital; P, parietal; PM, premaxilla; pmf, promaxillary fenestra; PO, postorbital; PRE, prearticular; Q, quadrate; QJ, quadratojugal; SA, surangular; soc, supraoccipital; SQ, squamosal.
Image and caption by Eddy & Clarke 2011 fig.2 (doi:10.1371/journal.pone.0017932.g002), Creative Commons Attribution