Eutheria
Taxa on This Page
- Adapisoriculidae X
- Bobolestes X
- Deccanolestes X
- Montanalestes X
The origin of Placental Mammals
As with amphibians and reptiles, our understanding of the evolutionary tree of mammals has undergone some major changes and paradigm shifts in the last half century. In part this is due to new discoveries, and in part to new methodologies.
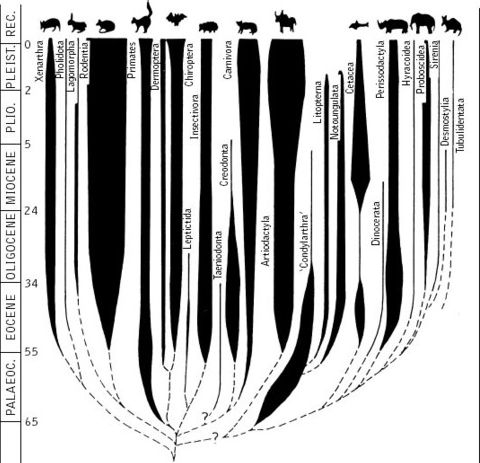
The classic understanding of mammals is based on vertebrate paleontologists such as Simpson and Romer who in the early to mid 20th century created the definitive approach that would be authoritative until at least the 1980s. This is shown by the diagram on the left, which is sometimes informerly called a romerogram, as it was Romer himself who popularised (but did not invent) such diagrams. Alternative names are spindle or bubble diagrams. As these terms indicate, these portray the evolution of a group in terms of a spindle or bubble or balloon of varying width. The width indicates either species, generic, or family diversity, or individual numerical abundance, or (rather unsystematically) both together. The vertical axis represents geological time. As shown in the late model diagram on the left - an earlier version, from Colbert 1969, accompanies our entry on Eutheria - mammals suddenly appear, as if out of nowhere, branching rapidly into the main groups (Linnaean orders). Although there are a few ancetsral or stem groups, such as Insectivora and Condylartha (this is made clearer in Colbert's diagram), the origin and inter-relationships of the various lineages remains mysterious.
Hennigian phylogenetic systematics (early cladistics), which became the central methodology in understanding reptile/amniote evolution, thanks to the revolutionary work of Gauthier and co-workers, becfore being supplemented and then replaced in the 90s and noughties by computer-generated statistical parsimony analysis (modern cladistics), was never central to Mammalian evolution. Instead, molecular phylogeny was the methodology of choice. This can be explained as due to the preponderance of extant lineages of mammals, whereas in the case of more ancient groups such as reptiles, amphibians, and early fish, most lineages are known solely from fossils.
As molecular sequencing has become more sophisticated and reliable, a consensus has emerged, strongly supporting new monophyletic clades as Afrotheria, Boreoeutheria, Euarchontoglires, Laurasiatheria and Xenarthra, for none of which is there any morphological evidence (synapomorphies) whatsoever. The most interesting thing is that the three main clades revealed through molecular phylogenomics, Afrotheria, Xenarthra, and Boreoeutheria [1] each are based on geographic origin. We will return to possible implications of this shortly.
Meanwhile older groups, such as the venerable Insectivora and Edentata, dissappeared entirely, going the way of Thecodontia and Captorhinomorpha, which fell victim to the cladistic revolution. Or rather, were reinterpreted as polyphyletic. Finally, there are a few fascinating new groups, most especially the Cetartiodactyla, where fossils, morphology, and molecules achieve a rare consensus
The following diagram, from Kriegs et al 2006, is representative of the new, and now standard, eutherian phylogeny:
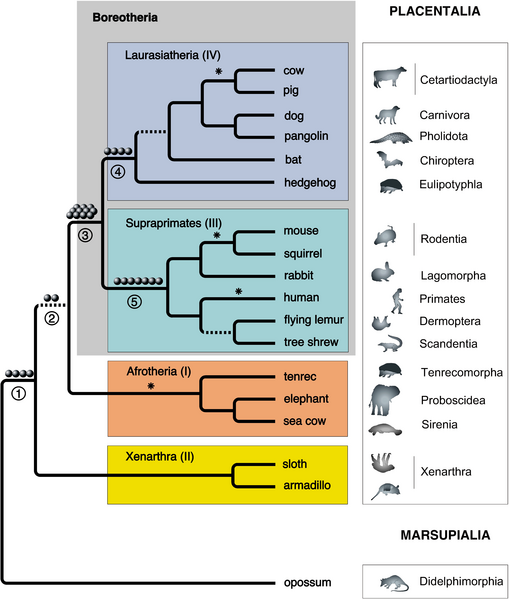
"Reconstruction of the placental mammalian (eutherian) evolutionary tree has undergone diverse revisions, and numerous aspects remain hotly debated. Initial hierarchical divisions based on morphology contained many misgroupings due to features that evolved independently by similar selection processes. Molecular analyses corrected many of these misgroupings and the superordinal hierarchy of placental mammals was recently assembled into four clades. However, long or rapid evolutionary periods, as well as directional mutation pressure, can produce molecular homoplasies, similar characteristics lacking common ancestors. Retroposed elements, by contrast, integrate randomly into genomes with negligible probabilities of the same element integrating independently into orthologous positions in different species. Thus, presence/absence analyses of these elements are a superior strategy for molecular systematics. By computationally scanning more than 160,000 chromosomal loci and judiciously selecting from only phylogenetically informative retroposons for experimental high-throughput PCR applications, we recovered 28 clear, independent monophyly markers that conclusively verify the earliest divergences in placental mammalian evolution. Using tests that take into account ancestral polymorphisms, multiple long interspersed elements and long terminal repeat element insertions provide highly significant evidence for the monophyletic clades Boreotheria (synonymous with Boreoeutheria), Supraprimates (synonymous with Euarchontoglires), and Laurasiatheria. More importantly, two retropositions provide new support for a prior scenario of early mammalian evolution that places the basal placental divergence between Xenarthra and Epitheria, the latter comprising all remaining placentals. Due to its virtually homoplasy-free nature, the analysis of retroposon presence/absence patterns avoids the pitfalls of other molecular methodologies and provides a rapid, unequivocal means for revealing the evolutionary history of organisms."
- diagram and text Kriegs et al 2006 - Creative Commons Attribution license.
Earlier findings in molecular phylogeny have thrown up some truly bizarre and unlikely groups, such as the Marsupionta hypothesis (monotremes + marsupials - Kirsch & Mayer 1998, now conclusively refuted Killian et al 2001, Kullberg et al 2008) or (in the case of reptiles) crown archosaurian turtles, which makes one wonder at the distorting influence of artifacts such as long branch attraction and molecular convergence. The field seems to be less problematic now, especially in the case of well sampled groups, and following the anomalous results of the 1990s and early 2000s, a strong consensus is now emerging. And whilst all trees may not be exactly like the one shown here (compare for example Murphy et al 2007 or Nishihara et al 2009) they are close enough that we can be confident regarding the status of the five major clades.
Although a strong critique of molecular phylogeny is incuded here, not all contributers to Palaeos necessarily feel this way (indeed, Palaeos would be a less interesting place were it not for the occasional disagreements and differences of perspectve between its contributing editors and compilers). For example, both morphology and molecules could be accommodated if we assume that the insectivora are neither monophyletic (morphology) or polyphyletic (molecules) but paraphyletic, an evolutionary grade that includes the ancestors and basal representatives of two of the major clades revealed through molecular phylogeny.
What has not yet emerged (and probably will not for some time) is an integral synthesis of (a) strato-phenetic or evolutionary systematics (for which it is hard to find any supporters now, despite the fact that it has never been disproved), both (b) the original Hennigian and (c) the more recent computiational-statistical versions of morphology based cladistics, (d) the little known field of stratocladists, and (e) molecular phylogeny used as one contrinuting methodology among many, rather than as now (due it being the current dominant paradigm) the sole approach.

Three phylogenetic hypotheses of the branching of major clades of placentals Tree 1, Exafroplacentalia (Afrotheria basal); tree 2, Epitheria (Xenarthra basal); and tree 3, Atlantogenata (Boreotheria basal). Diagram from Nishihara et al 2009 |
Acknowledging the current incompatibility of these various methodologies, we will continue our quixotic attempts at finding not a mere consensus, but the hint of a larger and more inclusive paradigm that include all these perspectives and methodologies.
Whilst there is, as mentioned, complete agreement from molecular phylogenists regarding the five major clades of placental mammals, two problems remain unresolved. These are (a) the branching order of the three major clades (Afrotheria, Boreoeutheria, and Xenarthra) and (b) the timing of the evolutionary radiation of eutherians; i.e. whether most Mesozoic eutharians (apart from a few obviously very primitive pre-Placental forms) preceeded or came after this branching point.
Taking the first of these, the branching order, support, and evidence, seem to be equally divided among the three options, although tree 2 is the only one which has even weak morphological support. Bininda-Emonds et al 2007 give a molecular clock date of around 101 mya for the three clades. Nishihara et al 2009 were not able to resolve which of the three possible origins were more likely. Retroposon analysis with mammalian genomic data revealed instead a surprisingly similar number of informative retroposon loci for each of hypotheses: 22 loci for the basal position for Afrotheria, 25 loci for Xenarthra, and 21 for Boreotheria, which shows the divergence of the crown placental common ancestor into the three lineages occurred nearly simultaneously.
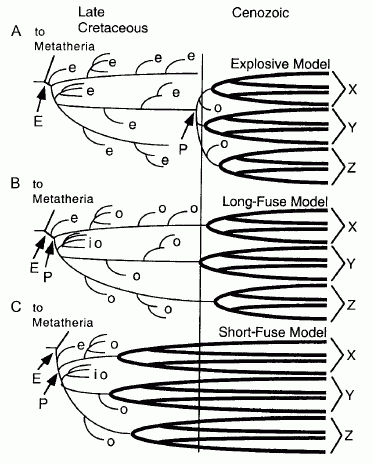
Three hypotheses of the timing of origin of extant placental mammal groups and the relationship of fossil Cretaceous taxa. The three hypotheses are (A) explosive, (B) long fuse, and (C) short fuse. Key: E, Eutheria; e, eutherian stem taxon; io, stem taxon to more than one ordinal crown group; o, ordinal stem taxon; P, Placentalia. Bold lines refer to crown groups. X,Y,Z, are extant placental orders and superorders. The vertical line marks the K-T Boundary. (From Archibald and Deutschman, 2001 via Rose 2006. A similar but simpler version of this diagram is presented in Gentili n.d. |
The second problem is not so easily resolved, if it can even be resolved at all. Here again there are three main hypotheses. the differences here are contradiction or incomgruence between molecular clock data and the fossil record. It seems that both cannot be correct (and hence, unlike the situation with strato-phenetic evolutionary systematics and morphology-based cladistics, which are complementary rather than contradictory, in this case there truly only can be one).
As shown by the diagram on the left, there are three hypotheses here, referred to as explosive, long fuse, and short fuse respectively. Of these, the first is based on fossils, and assumes a long separation between the first eutharian and the first placental, the latter two are based on the molecular phylogeny clock (and to some extent paleogeography), and has the eutharian and the placental origin close together (shown by the arrows pointing to the "E" and "P"). So essentially there's two alternatives, but one of these has two variations. In any case, if one of the members of the series seem out of place (i.e. explosive should go at the end, completing the comic book sequence), well, we just report the science, which is not to say we don't like to present our own take on things. Regarding which, let's turn the whole chart upside down and report the short fuse first.
The Short Fuse Model recently received a boost thanks to a popular paper by Bininda-Emonds et al 2007. It relies on molecular clock estimates while ignoring the fossil evidence (or, as regards crown placental mammals in the Cretaceous, the total lack therof). Assuming a constant molecular clock, all the extant placental mammals originated deep in the Cretacous and diverged long before the extinction of the dinosaurs. In other words mammalian diversity was already high even during the late Cretaceous. Bininda-Emonds et al 2007 argued on the basis of the molecular clock dating that per-lineage diversification rates barely changed across the cretaceous/Tertiary boundary. Instead there were two broad periods of evolutionary expansion among the mammals: an early event involving the origin of the currently recognized extant placental superorders and orders approximately 93 million years ago, and a more recent radiation of modern families throughout the Eocene and Oligocene epochs (re this see also PZ Myers blog post).
Unfortunately, this isn't what the fossil record tells us. Wheras only six species of shrew-like placentals are known from the late Cretaceous, by the earliest Paleocene, thirteen species, including nine archaic ungulates of three families, are known. By the middle Paleocene, there were 84 genera of mammals (mostly eutherians/placentals) in North America alone, and by the late Paleocene a hundred (Prothero 2006, p.55). The global diversity of mammalian families rose from 21 in the latest Cretaceous to 37 in the early Palaeocene, 86 in the late Palaeocene and 111 in the early Eocene Benton 2005 p.333.
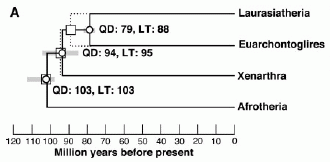 molecular phylogeny and clock by Murphy et al 2001. showing maximum likelihood molecular divergence estimates for the early radiation of placental mammals, estimated with the quartet-dating (QD) and linearized tree (LT ) methods. Open circles are median point estimates based on the former, open squares are point estimates based on the latter. Gray bars are range of 95% confidence intervals based on QD. molecular phylogeny and clock by Murphy et al 2001. showing maximum likelihood molecular divergence estimates for the early radiation of placental mammals, estimated with the quartet-dating (QD) and linearized tree (LT ) methods. Open circles are median point estimates based on the former, open squares are point estimates based on the latter. Gray bars are range of 95% confidence intervals based on QD.
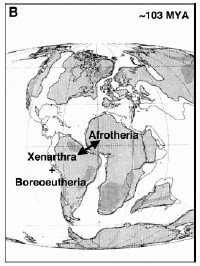 Paleogeographic map by Murphy et al 2001 shows how the separation of Africa and South America about 100 to 120 Mya, isolates the Afrotheria in Africa and the Exafroplacentalia (the clade that includes the common ancestor of Xenarthra and Boreoeutheria) in South America. Here the origin and early evolution of Eutharian mammals is anchored firmly in Gondwana. In a more recent paper however, Murphy et al 2007 reject the out of Africa hyposthesis in favour of the Atlantogenata hypothesis, which places the origin of the Eutheria in the northern continents Paleogeographic map by Murphy et al 2001 shows how the separation of Africa and South America about 100 to 120 Mya, isolates the Afrotheria in Africa and the Exafroplacentalia (the clade that includes the common ancestor of Xenarthra and Boreoeutheria) in South America. Here the origin and early evolution of Eutharian mammals is anchored firmly in Gondwana. In a more recent paper however, Murphy et al 2007 reject the out of Africa hyposthesis in favour of the Atlantogenata hypothesis, which places the origin of the Eutheria in the northern continents
|
The Long Fuse Model is something of a middle position. Although based on molecular data rather than fossils, it is very similar to the explosive model in taht it posits all the extant groups only appearing at the start of the Cenozoic. It still assumes a very early split between Afrotheria, Xenarthra, and Boreotheria (Murphy et al 2001), with the latter quickly further dividing into Euarchontoglires, Laurasiatheria (Bininda-Emonds et al 2007)). In this way, the three or four main Cenozoic clades originated deep in the Cretaceous, but passed through an extended lag, a "long fuse", before undergoing an evolutionary explosion. Hence there was a long period between the Cretaceous origins and the radiation of the crown groups at the start of the Cenozoic. A biogeographic scenario for the basal divergence among crown-group placental mammals, using the "long fuse" hypothesis, is presented by Murphy et al 2001, shown on the left. Again, it probably doesn't matter which of the three clades is more basal, since all would seem to have diverged more or less simultaneously. While in terms of retroposon analysis (Nishihara et al 2009) Xenarthra just pips the others at the post, which supports the preferred phylogeny of Kriegs et al. 2006 as against that of Murphy et al 2007, and also morphological data such as the epitheria hypothesis (although that has also been challenged), this is not a big enough difference to base a robust phylogeny upon, and the current consensus is basically a trifurcation of the three superorders of mammals (Churakov et al 2009, Nishihara et al 2009)
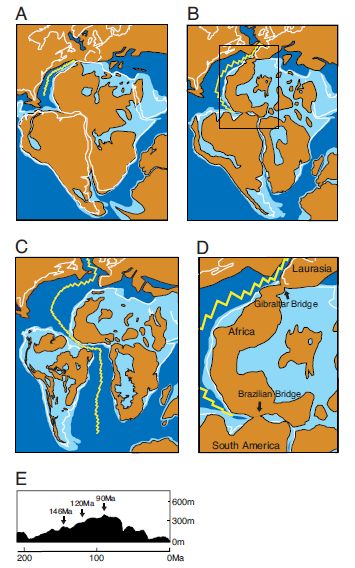
Paleogeographic scenario by Nishihara et al 2009. (A) Paleogeographic reconstruction of Pangea at 146 Ma, (B) at 120 Ma, and (C) at 90 Ma. (D) enlarged area of 120 Ma map (boxed area in B) showing two land bridges that connect three continents. Blue and light blue represent deep and shallow ocean depths, respectively. Brown areas represent land masses, the and the yellow jagged line is the mid-Atlantic ridge, respectively. Only deep ocean is shown for Laurasia because drilling data are currently unavailable. (E) Sea-level rise redulting in the fragmentation of Africa and South America. |
Although the standard molecular clock date for this speciation is a little over 100 mya (Murphy et al 2001 , Bininda-Emonds et al 2007), Nishihara et al 2009 suggest a slightly older timeframe with the divergence of Africa, South America, and Laurasia around 120 Ma. Although for several millions of years, these continents would have been separated by what amounts to little more than ocean straights. At various points there would be the opportunity for land bridges and archipeligos between adjacent continets can appear or conversely dissapear, continental drift would ensure their eventual mutual isolation. While it seems that the paleogeography is more persuasive than it is with the "sudden explosion" hypothesis, the latter is supported by evidence of a North-South American faunal interchange during the Paleocene (Pascual & Jaureguizar 1991), and the apparently total absense of any South American eutharians prior to that date (despite a strong record of Dryolestids (Rougieretal2011)). Apart from land bridges, small animals could still colonise islands and even continents through fortuitous "waif" dispersal, arriving for example by clinging to drift wood or other such circumstances (which is how rodents colonised Australia and monkeys South America).
Like the short fuse, the long fuse still suffers from the lack of evidence of any extant placental ordinal clades in the Cretaceous, or for that matter any Mesozoic placentals at all (Wible et al 2005, Archibald & Averianov 2007, Wible et al 2009, Giallombardo 2009, and Goswami et al 2011. It might be plausibly suggested that Cretaceous eutharians are stem-group representatives of one of the three or four major clades, and the long fuse model supported in this way, although this is still contrary to current cladistic and morphological analyses. The implications for the short fuse model are more severe. It is hard to imagine that the late Cretaceous was populated by such a rich diversity of placental mammals, many from extant clades, without a single one ever being fossilised.
A compromise between discrepancies between the fossil record and molecular clock data may be explained by the Garden of Eden hypothesis, with a long period of cryptic evolution in southern continents, but more evidence of numerous lineages of extant eutherians originating in the Southern Hemisphere are needed to corroborate such a model Gentili n.d.. Gentili suggests the Australian Ausktribosphenidae as a representative early taxon, but this is now known to lie outside the main eutharian clade. Moreover, the same lack of evidence of late Cretaceous eutharians in the otherwise well represented and sampled South American fossil faunas also argues against the "Garden of Eden" hypothesis.
This leaves us with the Explosive Model. As the name indicatse, this assumes a long period in which nothing much happens (as with the long fuse model), followed by a sudden explosive adaptive radiation. The Explosive Model is based on paleontological, and more recently cladistic-morphological evidence (see diagram below right). It reveals Cenozoic mammalian diversity as the product of an opportunistic radiation of species after the end-Cretacous extinction of the dinosaurs (or to be cladistically pedantic, of non-avian dinosaurs) , 65 million years ago. It is supposed that at this time there was a rapid origination and diversification of extant placental orders. Accordig to this hypothesis, all Mesozoic eutharians are therefore stem-eutharia, and are placed outside the placental crown group (the clade that includes the common ancestor of all living mammals, and all its descendents)
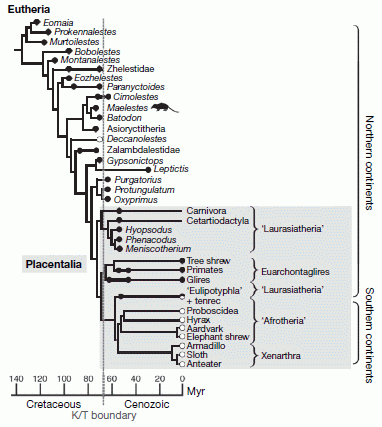
Diagram from Wible et al. 2007, fig.3 showing the of Maelestes (silhouette ) to other eutherians, as well as crown group Placentals. The cladogram shows the strict consensus of three most parsimonious trees (2,296 steps). The timescale is shown at the bottom, taxa marked stratigraphically. Filled circle taxa, Laurasian (northern continents); open circle taxa,Gondwanan (southern continents). Circles and circles with thicker black lines indicate temporal occurrence of studied specimens. Broken line, K/T boundary at 65 million years. The grey box delimits Placentalia. Wible et al. 2009 provides a more detailed analysis of basically the same tree. |
The explosive model and laurasian origin (Atlantogenata hypothesis) is presented in this diagram from Wible et al. 2007, based on study of a newly discovered Cimolestid taxon, Maelestes gobiensis. The cladogram places all Mesozoic eutharians, without exception, outside the placental crown group, shown by the grey box at the bottom right. Here Placentalia is defined not physiologically or apomorphically but in terms of crown groups - the latest common ancestor of all extant placentals, and all its descendents. A similar cladistic definition is made for Amniota and sometimes Tetrapoda. By this definition Placentalia - and hence the ancestry of all current groups - originates at or near the K/T boundary in Laurasia.
Significantly, the only Mesozoic Gondwanan taxon shown, Deccanolestes from the latest Cretaceous of India, has since been argued to be a basal Euarchontan (Boyer et al 2010, Smith et al 2010) rather than, as suggested by Wible et al. 2007, an independent southern migration of stem eutherians. If Deccanolestes is a crown placental, this also supports a southern origin as opposed to a northern one (Prasad 2009 Boyer et al 2010). At the time India was an island continent, like Australia today, and hence the fauna would have been quite isolated. While one can assume rafting across island archipeligos, clinging to driftwood and so on (Placental rodents reached Australia in this way) it still seems hard to imagine indigenous Paleocene mammals being supplanted in this way. Animals evolving on large continents would also seem to have the advantage over endemic island or small continents; which is why most of the Cenozoic has been dominated by boreotheres, with only the Proboscideans venturing out of Africa as global megaherbivors. But the microcontinent problem can be circumvented if we assume that this was an early group of stem-euarchontoglires that became isolated with the separation of India, and hence continued evolving in ther own way, much like monotremes and marsupials in Australia during the Cenozoic
It might be suggested that a problem with Wible et al. 2007's cladogram is that the predominance of Laurasian (and specifically Mongolian and American) Mesozoic forms would clearly be expected to skew the results, hence Boreoeutheria would naturally appear as the most basal placental clade. And the step-wise arrangement of taxa may be the result of convergence; most Mesozoic eutharians would have had a very similar morphology and lifestyle. A more recent cladistic analysis by Goswami et al 2011 supports a robust relationship between Deccanolestes and Paleocene adapisoriculids, but does not show any close affinity between these taxa and crown placentals, as well as previously unrecognised interchange between Europe, India and Africa. This reinforces the findings of Wible et al. 2007 and Wible et al. 2009, reinforcing the sizeable gap between molecular and fossil divergence time estimates for the placental mammal radiation.
Summing up, fossil evidence and the absense of crown Placental groups provides a strong argument against both the "long fuse" and "Garden of Eden" hypotheses, and (adding to the incongruency with the Pal;eocene fossil record) an even stronger one against the "short fuse" model. The unavoidable implication is that molecular clocks are notoriously unreliable, and that evolutionary rates would vary, depending on circumstances. This also argues against Motoo Kimura's Neutral theory of evolution, according to which most mutations are selectively neutral and accumulate at a fairly constant rate. If instead natural selection is a determining factor, or, perhaps most plausibly, the input of each varies according to circumstancxe and opportunity fora daptive radiation, rapid adaptive radiation would give an appearance (in molecular clock dating) that a group is older and diverged earlier than it actually did. Hence the great Cretaceous placental radiation is simply an artifact of rapid Paleocene and early Eocene evolution. For this reason, and until evidence suggests otherwise, we have decided to support the "Sudden Explosion" hypothesis of placental evolution. An added, rather startling, implication here is that even many Paleocene mammals may turn out not be placentals at all! MAK111208
Notes
[1] for some obscure reason this spelling seems to be more popular and widely used than the altogether more sensible "Boreotheria", as if six syllables wasn't enough), are geographically based ,of Africa, South America, and Laurasia respectively. Gar be it for the present author to interfere with such a venerable tradition
Descriptions
Adapisoriculidae Van Valen 1967 : Adapisoriculus, Afrodon, Bustylus, Deccanolestes, Garatherium
Range: Late Cretaceous to Late Paleocene, Europe, Africa, and India
Phylogeny: Eutheria ::: (Zhelestidae + (Asioryctitheria + Cimolestidae) + (Zalambdalestidae + (Leptictida + Placentalia))) + *.
Comments: Insectivore-like mammals, about 15 cm long, with a tail of equal length. They were probably nocturnal, eating insects and fruits. They were once thought to be related to Hedgehogs (Erinaceidae), because of their similar dentition, then considered basal Euarchontans (Smith et al 2010, Boyer et al 2010). A new study by Goswami et al 2011 supports a robust relationship between Deccanolestes and Paleocene adapisoriculids, placing the group in a much more basal position than previously thought, and strengthening hypotheses that scansoriality and arboreality were important early in eutherian evolution. The implication is that a faunal exchange occurred between India, Africa, and Europe in the Late Cretaceous and Early Paleocene, and implies a previously unrecognized ~30 to 45 Myr ghost lineage for these Gondwanan eutherians.
Deccanolestes is known from two species, Deccanolestes hislopi (Prasad and Sahni, 1988) Deccanolestes robustus (Prasad, Jaeger, Sahni, Gheerbrandt and Khajuria, 1994) from the Late Cretaceous (Maastrichtian) Intertrappean Beds of Andhra Pradesh, India. The former is known from a few teeth, the latter from a single molar. Later Deccanolestes narmadensis Prasad et al. 2010 was added. With such scanty remains it's amazing anything can be known about these little creatures at all. The equally poorly known Sahnitherium comes from the same locality. Deccanolestes has been previously referred to Palaeoryctidae, but this assignment was rejected (Rana & Wilson 2003, p.331). More recent findings including postcrania resulted in reinterpretationof Deccanolestes as a basal euarchontan. The humerus resembles those of euarchontans in possessing a laterally keeled ulnar trochlea, a distinct zona conoidea, and a spherical capitulum (Boyer et al 2010), but these may simply be convergent features consistent with an arboreal/scansorial lifestyle. A basal Euarchontan assignment supports the "Long Fuse" hypothesis (Prasad 2009), with the Indian subcontinent, Eurasia, and Africa are more likely places of origin for Euarchonta than is North America (Boyer et al 2010). Further study however reinterpreted it as the earliest known Adapisoriculid (Smith et al 2010, Goswami et al 2011), and it is now considered a stem-eutharian (Wible et al. 2007, Wible et al. 2009, Goswami et al 2011).
Links: Mesozoic Mammals, Wikipedia Wikipedia MAK111208