
| Therapsida | ||
| The Vertebrates | Theriodontia |
| Vertebrates Home | Vertebrate | Vertebrate |
Abbreviated Dendrogram
Synapsida
│
└─Therapsida
├─Biarmosuchia
└─┬─Dinocephalia
│ ├─Anteosauria
│ └─Tapinocephalia
└─┬─Anomodontia
│ ├─Venyukovioidea
│ └─Dicynodontia
└─Theriodontia
├─Gorgonopsia
└─Eutheriodontia
├─Therocephalia
└─Cynodontia |
Contents
Index |
The third great therapsid taxon, the Theriodontia "beast tooth", referring to the mammal-like teeth), include the mammals. Even their most basal members are the most mammal-like of the Therapsida. The theriodonts included both carnivores and herbivores, and various lineages correlated to late Cenozoic wolves, weasels, otters, rodents, and shrews. Most diverse and successful during the Late Permian and Early Triassic, they were increasingly overwelmed by the archosaurian radiation of the later Triassic. During the latest Triassic (late Norian to Rhaetic) at least one line of eutheriodonts evolved into mammals. Even so, one family, the rodent-like tritylodontids continued right into the Late Jurassic (Oxfordian).
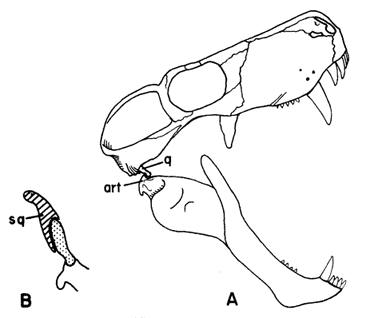 |
| The theriodont specialization of the masticatory apparatus illustrated by the gorgonopsian Dixeya. In A, the jaws are widely opened; the articular (art) tightly grips the head of the small quadrate (q). The lower incisors pass entirely behind the upper incisor~ when the jaws are closed. B shows a diagrammatic cross section through the squamosal (sq) showing movable quadrate in squamosal recess. (A modified from Parrington.) |
The theriodonts evolved a feeding mechanism much more successful than that of the Dinocephalia. The quadrate bone in the jaw was reduced in size. When the jaws are widely opened (drawing, right) the articular bone (art in the diagram) toghtly grips the head of the small quadrate (q). The lower incisors pass entirely behind the upper incisors when the jaws are closed. The development of versatile jaw movements and a more precise type of dental occlusion also played a critical role in preadapting the elements of the theriodont articular complex for becoming the ossicles of the mammalian middle ear.
The small quadrate may have been loosely attached to the skull, probably by a fibrous pad in life; set in a squamosal recess (B) in the diagram above. The upper end occupies a hollow in the anterior face of the squamosal which also buttressed it from behind. Thus, as Hopson 1969) suggests, the quadrate was probably capable of a small amount of fore-aft movement at its lower end. Nevertheless the joint between the quadrate and articular was a tight one, with the articular firmly gripping the quadrate condyle, allowing only a hinge (and some transverse sliding) movement at the jaw joint, but no fore-aft sliding.
The reduced and movable quadrate was a key theriodont adaptation. In large carnivores with very long canines, such as the gorgonopsians and some therocephalians, a wide gape was necessary, and so a secure attachment of the lower jaw is required. This same adaptation is also found in mammalian carnivores. The theriodont quadrate also allowed the anterior (front) surfaces of the lower teeth to contact and shear against the posterior (rear) surfaces of the upper teeth as the jaws are closed. These teeth, which were sometimes serrated in gorgonopsian and other groups, could then function as very effective cutting instruments for dismembering a carcass or biting out chunks of flesh from it. In advanced herbivorous cynodonts, like the tritylodonts, the ability to move the lower jaws in a fore-aft direction was ex- ploited for the purpose of cutting up plant material by the cheek dentition.
 The theriodonts fall into three distinct groups, the Gorgonopsia; the Therocephalia, and the Cynodontia. A fourth group, the Bauriamorpha, are now generally included with the Therocephalia. The Gorgonopsia were the "sabre tooth tigers" of the Permian, medium-sized to large therapsids equiped with huge canine teeth, used to bring down large prey. These theriodonts, typified by animals like the dog-sized Lycaenops (below) and the bear-sized Inostrancevia, show some features that were further developed in the later theriodonts. Thus the dentary bone in the jaw was large, but not as large as in Cynognathus. In mammals, the jaw consists only of the dentary. The teeth, although differentiated, were not highly specialized. There was as yet no hard secondary palate (bony roof of the mouth, enabling the animal to eat and breath at the same time). The occipital condyle, which connects the skull to the backbone, was single.
The theriodonts fall into three distinct groups, the Gorgonopsia; the Therocephalia, and the Cynodontia. A fourth group, the Bauriamorpha, are now generally included with the Therocephalia. The Gorgonopsia were the "sabre tooth tigers" of the Permian, medium-sized to large therapsids equiped with huge canine teeth, used to bring down large prey. These theriodonts, typified by animals like the dog-sized Lycaenops (below) and the bear-sized Inostrancevia, show some features that were further developed in the later theriodonts. Thus the dentary bone in the jaw was large, but not as large as in Cynognathus. In mammals, the jaw consists only of the dentary. The teeth, although differentiated, were not highly specialized. There was as yet no hard secondary palate (bony roof of the mouth, enabling the animal to eat and breath at the same time). The occipital condyle, which connects the skull to the backbone, was single.
These were efficient, very capable hunters, the dominant predators of the Late Permian terrestrial ecosystem. They may have hunted singly or in packs, and probably preyed on large, plant-eating animals like Pareiasaurs, Tapinocephalians, and large Dicynodonts. The canines were particularly long, giving the mouth a distinctly "sabre-toothed" appearance, and the front part of the skull was deeper than normal to accommodate their roots.
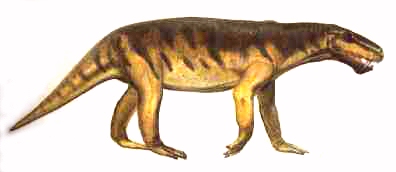 A medium-sized form like Lycaenops, represented by several species from the Late Permian (Cistecephalus and Daptocephalus zones) of South Africa, averaged about a meter in length. It was a lightly built carnivore, with long running legs, that almost certainly fed on the abundant dicynodont herbivores that shared its environment. Larger forms, like Dinogorgon and Inostrancevia, grew to be lion to bear sized, and prayed upon the ox- and hippo-like megaherbivores like the Pareiasaurs and the biggest Dicynodonts, both of which attained weights of a tonne. Gorgonopsids and Pareiasaurs may have formed a co-adaptive relationship (like Pleistocene megaherbivores and sabre-tooth cats), and significantly both became extinct at the same time (at the end of the epoch)
A medium-sized form like Lycaenops, represented by several species from the Late Permian (Cistecephalus and Daptocephalus zones) of South Africa, averaged about a meter in length. It was a lightly built carnivore, with long running legs, that almost certainly fed on the abundant dicynodont herbivores that shared its environment. Larger forms, like Dinogorgon and Inostrancevia, grew to be lion to bear sized, and prayed upon the ox- and hippo-like megaherbivores like the Pareiasaurs and the biggest Dicynodonts, both of which attained weights of a tonne. Gorgonopsids and Pareiasaurs may have formed a co-adaptive relationship (like Pleistocene megaherbivores and sabre-tooth cats), and significantly both became extinct at the same time (at the end of the epoch)
The Gorgonopsia are generally considered the most primitive members of the Theriodontia. It is also conceivable that the characters which they share with more mammal-like therapsids could perhaps be explained as convergences (parallel evolution), rather than as shared traits inherited from a common ancestor. The Gorgonopsia have thus been grouped by Boonstra with the Dinocephalia and other very primitive forms such as the Bairmosuchia, under the name "Alphatherapsida". However there is no certain evidence for a close relationship between the Gorgonopsia and Dinocephalia, especially since many primitive forms (e.g., the Ictidorhinidae) previously considered gorgonopsians are now generally accepted not to belong to this group at all. The most likely hypothesis at present is that the Gorgonopsia represent a distinct lineage of theriodonts. MAK000718.
The Therocephalia are at least as ancient as the Gorgonopsians, which they resemble, and in fact outlasted, making it through to the Early Triassic period. The earlier therocephalians were in many respects as primitive as the gorgonopsians, but they did show certain advances such as the enlargement of the temporal opening (for a lighter skull and mopre efficent muscle attachment) and reduction of the phalanges (finger and toe bones) to the mammalian formula. The later therocephalians included the advanced bauriamorphs, which carried some theriodont characters to a high degree of specialization. For instance, in Bauria there was no bar of bone separating the orbit from the temporal opening; a condition typical for primitive mammals. These and other advanced features led to the long-held opinion, now rejected, that the Ictidosaurs and even some early mammals arose from a bauriamorph stem. Rather, the situation seems to be that mammalian characteristics evolved in parallel among a number of different groups, and this makes the therapsid phylogeny a particularly tricky subject. [see for example Luo et al. (2002))
The fragmentary nature of some of these fossils, and the enthusiasm of early workers in the field to give each new scrap of bone a different species, genus, or even family designation, has led to a number of false taxa. It is not surprising that several therocephalian groups, like the Lycosuchidae and the Scaleposauridae, have turned out to be artificial, the former based on skulls which retain an extra set of canines (i.e. when the animal died one pair was in the process of replacing the other), and the latter based on mostly juvenile characteristics. MAK000907.
To be added...
from the Middle Permian.
Phylogeny: Neotherapsida: Anomodontia + *: Gorgonopsia + Eutheriodontia.
$ Skull flat with wide muzzle (dorsal surface of nasals horizontal) [RS01]; dorsal process of premaxilla shortened; $ skull roof narrow between tempral fenestrae (narrower than interorbital width) [RS01]; quadrate and quadratojugal shortened and lie in a pocket of the squamosal; $ dentary has coronoid process, providing longer lever arm and more area for insertion of jaw adductor and temporalis (in later forms) muscles [RS01]; post-dentary bones reduced; dentary canines longer in relation to maxillary canines.
Therapsida; Evolution of Mammals; dinosaurs-inostrancevia; Theriodontia; Annatherapsidus; Lecture 03 - Cont. Drift.
Comments: Primitive forms were small and rather rodent-like carnivores and insectivores. This is the approximate place on the mammal road that features like fur and elevated metabolic rates are thought to have first occurred.
Rubidge & Sidor (2001) [RS01].
ATW020206.
checked ATW030505; last modified MAK091111
