
Tetraceratops - life reconstruction
artwork by Dmitry Bogdanov - Wikipedia
| Therapsida | ||
| The Vertebrates | Therapsida |
| Vertebrates Home | Vertebrate | Vertebrate |
Abbreviated Dendrogram
Synapsida
│
└─Therapsida
├─Tetraceratops
├─Raranimus
└─┬─?Phthinosuchidae
├─Biarmosuchia
└─┬─Dinocephalia
└─┬─Anomodontia
│ ├─Venyukovioidea
│ └─Dicynodontia
└─Theriodontia
├─Gorgonopsia
└─┬─Therocephalia
└─Cynodontia |
Contents
Index |
|
|
Tetraceratops - life reconstruction artwork by Dmitry Bogdanov - Wikipedia |
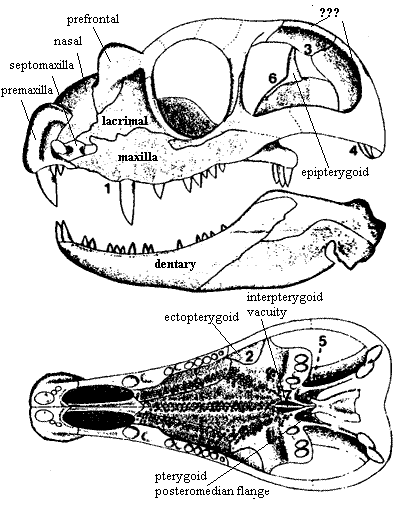
The generic name of Tetraceratops insignis Matthew 1908 means "four-horned face," although it has at least six protruberances, none of which are horns. That original misnomer is symptomatic of the confusion surrounding this animal. The fossil, a 95 mm unque skull, was originally recovered from the Early Permian Arroyo Formation (Clear Fork Group) of Baylor County, Texas. It was initially identified as an eothyrid pelycosaur of uncertain relationships. The only known specimen was embeded in a particularly recalcitrant rock matrix and was very difficult to prepare.
In 1996, Michel Lauren and Robert Reisz published an important paper on Tetraceratops, based on additional preparation. Laurin & Reisz (1996). Many of their general anatomical findings are discussed on the previous page. These workers argued that this strange little animal was actually a connecting link between the basal syanapsid "pelycosaurs" and the therapsids.
At least three pairs of horns are present on this animal, one on each premaxilla, prefrontal, and angular bones of the skull. These give the animal a superficial similarity to the primitive therapsid Burnetia mirabilis, a biarmosuchian. However, this does not necessarily indicate close relationships, because the horns are not located on the same bones. It is interesting to coinsder that similiar hornlets are found on a number of different (and only distantly related) theropod dinosaurs - Dilophosaurus, Cryolophosaurus, Allosaurus, Carnotaurus, Tyrannosaurus, etc). It is not unlikely that in all these instances the horns may have been brightly coloured and served as instruments of intra-specific display.
Laurin & Reisz determined that Tetraceratops and the therapsids form a monophyletic group because they share the following seven derived features (synapomorphies) which appear in the primitive condition in sphenacodontines (see diagram at right):
The presence of a number of diagnostic therapsid features cannot be tested in Tetraceratops because of the fragmentary nature of the specimen. However, all other therapsids would seem to form a monophyletic group excluding Tetraceratops because they share the following derived characters that are not present in Tetraceratops and pelycosaurs:
This combination of primitive and advanced features showed that Tetraceratops is intermediate between pelycosaurs and all previously known therapsids, an could serve as pretty good ancestral type (or "sister taxon" in cladistic jargon) to the later Therapsida.
More recently, however, Conrad & Sidor (2001) again reviewed this specimen and concluded that Tetraceratops was neither an eothyrid nor a therapsid. Rather, it was a sphenacodontid. Their detailed findings are mentioned on the previous page. In essence, they assert that the skull had been sheared over time. as a result, the entire arch of the posterior jaw was displaced upwards. As a result, what appeared to be the skull roof above the temporal fenestra was actually bits and pieces of the zygomatic (jaw) arch. This actually affects only character #3 above. However, this was enough to change the shape of the cladogram in a subtle, but significant way, shifting Tetraceratops onto a dead-end side track, the Sphenacodontidae (like Dimetrodon), rather than being on the main line to the Therapsida. However, whether Tetraceratops is a therapsid or just a close cousin, it is plainly a reasonably good guide to the sorts of forms which connect the "pelycosaurs" with the Therapsida.
It is quite likely that proto-therapsids such as Tetraceratops (and many others which never became fossilised) evolved in an upland environment where they were not easily fossilised, away from the swamps and deltas frequented by the Pelycosaurs (the only upland Pelycosaur lineages were the Caeseidae and, most certainly, their eothyrid ancestors). This early Permian upland evolution proceeded parallel to the Permocarboniferous lowland fauna, and probably originally derived from reptiles that adapted to dry habitats during the long Kazimovian arid period. It was from this upland evolution, via intermediate forms like Tetraceratops, that the great therapsid evolutionary radiation emerged.
MAK000802, ATW020518.
Overworked author's note: the following is from Wikipedia (quoted here in toto because it is so well-written and there are so many pages to get through...) - content mostly by Smokeybjb.
MAK091113
 |
Raranimus artwork by Dmitry Bogdanov - Wikipedia |
Raranimus is an extinct genus of therapsid of the Middle Permian. It was described in 2009 from a partial skull found in 1998 from the Dashankou locality of the Xidagou Formation, outcropping in the Qilian Mountains of Gansu, China. The genus is the most basal known member of the paraphyletic order Therapsida, from which the class Mammalia is a descendant taxon. (Liu, Rubidge, & Li 2009)
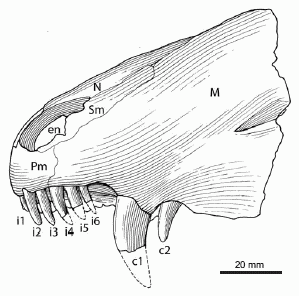 |
Partial skull of Raranimus dashankouensis (drawing of holotype) left lateral view. Abbreviations: c1–2, canine 1–2; ch, choana; en, external naris; i1–6, incisor 1–6; M, maxilla; N, nasal; p1–3, postcanine 1–3; Pf, prefrontal; Pl, palatine; Pm, premaxilla; Pt, pterygoid; rc, replacement canine; ri, replacement incisor; Sm, septomaxilla; sp, small precanine maxillary tooth; V, vomer. From Liu, Rubidge, & Li 2009 p.395. |
Raranimus shares a number of features with later therapsids and ancestral sphenacodontids. The skull consists of a well preserved rostrum. The teeth suggest a carnivorous lifestyle for Raranimus, as the incisors are recurved and the second canines are serrated on their posterior edges. The incisors are morphologically similar to those seen in more derived theriodonts. The presence of two linguo-labially compressed canines is a diagnostic feature of Raranimus. The presence of two functional canines is characteristic of sphenacodontids, and this condition is seen in no other therapsid other than Rananimus. However, the slender, compressed shape of these canines is a derived characteristic of therapsids, with the canines of similarly sized sphenacodontids being more massively built. The precanines are small and anteriorly serrated, similar to what is seen in the synapsids Dimetrodon and Tetraceratops.(Romer & Price 1940, Laurin & Reisz 1996]
In the palate region of the skull, the anterior process of the vomer ventrally overlies the premaxilla at the anterior margin of the choana. This overlap is also seen in dinocephalians. However, unlike any other therapsid, the choanae are short and extend only from the level of the fourth incisor back to the first canine.Liu, Rubidge, & Li 2009
According to a phylogenetic analysis conducted along with its initial description, Raranimus is considered to be the basalmost therapsid. Liu, Rubidge, & Li 2009 There has been some controversy as to whether or not Tetraceratops is a therapsid or a more basal pelycosaur. If Tetraceratops is a therapsid, as has recently been proposed, it would be the oldest and most basal one known, surpassing Raranimus in age by several million years.Laurin & Reisz 1996. However, later studies have questioned the placement of Tetraceratops within Therapsida, and the 2009 phylogenetic analysis using Raranimus places the genus outside of the clade. (Liu, Rubidge, & Li 2009, Sidor & Hopson 1998, Conrad & Sidor 2001]
Raranimus occurs in strata that were deposited during the early Roadian stage of the Middle Permian. [The hypothesized age for this locality is supported by the presence of the dissorophoid temnospondyl Anakamacops, the bolosaurid Belebey, and the basal therapsids Biseridens, Sinophoneus, and Stenocybus.] The sphenacodontids were most diverse before the Roadian in the Early Permian, yet therapsids did not appear as a diverse group until near the Roadian-Wordian boundary. This has left a morphological and temporal gap in the fossil record during which the origin of therapsids must have occurred. (Abdala et al 2008) This gap has been called "Olson's Gap".[Lucas 2004, Ivakhnenko 2005
With the general absence of therapsid remains found from strata deposited during Olson's Gap, different hypotheses have developed in order to explain the group's origins and initial diversification. One theory suggests that therapsids diversified quickly through rapid apomorphy accumulation sometime during the gap, while the other proposes that therapsids evolved gradually over the course of up to 35 Ma. (Abdala et al 2008, Kemp 2006] Only recently have remains of basal therapsids such as Raranimus been found from China that occur during Olson's Gap. Other therapsids that are known to have existed during the gap include Sinophoneus and Stenocybus.[Cheng & Li 1997]
 |
Titanophoneus potens courtesy mathematical.com. |
mammals > sphenacodontids [LR96].
From the Early? Permian [RS01].
Sphenacodontia: Sphenacodontidae + * : Tetraceratops + (Raranimus + (Phthinosuchidae + (Biarmosuchia + (Eotitanosuchia + Eutherapsida)))).
Large head with relatively rigid skull; $ long dorsal process of premaxilla [B&S, H&P] [RS01]; $ septomaxilla with posterodorsal facial process separating nasal & maxilla [B&S, H&P, contra LR96]; $ maxilla enlarged, separating nasal & lacrimal [B&S, H&P]; $ maxilla contacts prefrontal (except Tetraceratops) [LR96] [RS01]; $ enlarged temporal fenestra [B&S, H&B] [RS01]; $ upper margin of fenestra forms concave, ventrolaterally facing surface (for origin of jaw adductors?) [LR96]; $ pineal foramen raised on prominent boss [B&S, H&P] [RS01]; $ supratemporal absent [B&S, H&P] [RS01]; $ squamosal with groove on posterior [B&S, H&P]; $ quadrate reduced [LR96]; $ braincase firmly sutured to back of dermal skull roof [LR96]; dentary anteriorly expanded [RS01]; anterior coronoid absent [RS01]; $ posterior coronoid ventrally shifted & fails to form dorsal margin of jaw in medial view [RS01]; $ reflected lamina of angular bone notched dorsally [B&S, H&P] [RS01]; $ reflected lamina with pattern of ridges on lateral surface; $ vomer expanded transversely between choanae [B&S, H&P]; vomer with concave ventral surface; at least partial secondary palate; $ posteromedian flange of pterygoid present behind the interpterygoid vacuity [LR96]; $ pterygoid short & palatines meet at midline (except Tetraceratops) [LR96]; $ interpterygoid vacuity shortened [B&S, H&P, LR96]; basicranial articulation fused [RS01]; $ parasphenoid ventral plate ridged & lacks central groove (except Tetraceratops) [LR96]; $ precanine teeth absent; $ premaxillary teeth of equal size (except Tetraceratops) [LR96]; enlarged canine dentition [RS01]; $ upper canine length increased [B&S] $ no more than twelve upper postcanine teeth [LR96]; $ teeth absent from ectopterygoid [LR96]; $ palatal teeth restricted to raised denticle fields (except Tetraceratops) [LR96]; heavy body; $ intercentra absent from trunk [RS01]; more than 3 sacral vertebrae; tail reduced; limb girdles reduced and more mobile; limbs more slender, but fairly short; limbs held more vertically; $ scapular blade slender [RS01]; $ pectoral glenoid deepened & rounded [RS01]; $ humeral head rounded, not screw-shaped [RS01]; expanded iliac blade; $ acetabulum deepened & rounded [RS01]; $ inturned head of femur [RS01]; general reduction of ventral components (e.g. clavicle -- indicative of more upright posture); inflected femoral head [RS01]; development of greater trochanter on femur (with iliac blade, indicates gluteal hindlimb musculature); phalanges shorter (used as lever, rather than holdfast); possibly had hair; increased metabolic rate probable; found in terrestrial habitats with nearly world-wide distribution.
There are a remarkable number of good sites: Thrinaxodon; Paleontology and Geology Glossary: T; Therapsida; Therapsida; Untitled7; Mammal-like reptiles.
Battail & Surkov (2000) [B&S]; Hopson & Barghusen (1986) [H&B]; Laurin & Reisz (1996) [LR96]; Rubidge & Sidor (2001) [RS01].
ATW020206.
T. insignis Mathew, 1908.

Early Permian of North America.
Therapsida : + (Raranimus + (Phthinosuchidae + (Biarmosuchia + (Eotitanosuchia + Eutherapsida)))) + *.
Skull high & short; $ first premaxillary tooth much larger than others [LR96]; upper tooth row slightly convex; 1 small precaniniform; $ long diastema on maxilla anterior to caniniform [LR96]; maxilla with broad alveolar shelf; postcaniniform teeth with sharp, curved crowns; $ very large teeth on transverse flange of pterygoid [LR96]; ectopterygoid teeth absent; dentary largest jaw element; $ dentary ends ventral to coronoid process [LR96]; angular with reflected lamina (contra, [CS01]); posteroventral flange of angular may have had additional horn; surangular forms coronoid; surangular with prominent knob for articular; choana long, narrow, bordered medially by vomers; palatines ling & narrow with medial denticle field; ectopterygoid small & triangular; posteromedian flange of pterygoid present; pterygoids with medial and lateromedial denticle rows; ventral plate of epipterygoid (?) reduced; ventral plate of epipterygoid excluded from basicranial articulation; interpterygoid vacuity shortened; pterygoids meet medially anterior & posteriorly to vacuities; basipterygoid articulation present (not fused); $ parasphenoid ventral plate narrow [LR96], with large medial depression; parasphenoid fused with basisphenoid; premaxilla very deep; septomaxilla small (!? also described as "massive" in same paper) with no facial process; maxilla contributes to narial margin; $ bony, possibly horn-bearing, tuberosities on premaxilla, prefrontal and angular [LR96]; tuberosities covered with fine ridges laterally & small foramina dorsally (=distally); maxilla does not contact nasal; lacrimal contacts nares briefly; nasal small, with (probable) median ridge; large orbital contribution of the lacrimal [LR96]; prefrontal with large tuberosity (horn); deep posteroventral excavation of prefrontal; long thin ventral process on orbital rim under lacrimal; $ suborbital process of jugal very narrow [LR96], terminating in blunt wedge in maxilla; postorbital ramus of jugal overlaps postorbital extensively (but not "twisted" as in later forms); postorbital overlaps & firmly sutured to squamosal, excluding parietal from temporal fenestra (questioned by [CS01]); broad, concave shelf of squamosal on upper, internal margin of temporal fenestra (origin of jaw adductors); wide tabular overlapping occipital portion of squamosal ([CS01] assert that [LR96]'s tabular is actually a fragment of the squamosal); braincase attached firmly to cheek; $ paroccipital process (?) narrow [LR96] & cylindrical, attached to tabular and squamosal; quadrate reduced; quadrate with two clearly separated condyles of equal size.
Despite the name, Tetraceratops had at least 6 horns. Only 4 were exposed at the time it was named.
[CS01] argue convincingly that this is not a therapsid.
[LR96] is a paradigm of what a descriptive paper should be. If everyone wrote like this, the professional literature would be half as long and twice as useful. Highly recommended.
[CS01] argue that much of the skull roof area is actually the zygomatic arch. Based on a revised parsimony analysis, they assert that Tetraceratops is a garden-variety sphenacodontid.
New Page 1; The osteology and relationships of Tetraceratops insignis, the ...; The First Therapsids; Artist- Brad McFeeters [The Dinosauricon]
Conrad & Sidor (2001) [CS01]; Laurin & Reisz (1996) [LR96].
ATW031019.
R. dashankouensis Liu, Rubidge, & Li 2009.
Roadian (Early Middle Permian) of China. (Xidagou Formation, Dashankou, Gansu, China)
Therapsida : + (Biarmosuchia + (Eotitanosuchia + Eutherapsida)))) + *.
Plesiomorphic, short choana; long facial process of septomaxilla; one precanine and two functional linguo-labially compressed canines on maxilla; six incisors.
complete skull estimated to exceed 16 cm.
checked ATW040305; last modified MAK091114, 120127
{kind=link}
{kind=link}