Cynognathus. Early Triassic of South Africa.
From Cox et al. 1988).
| Therapsida | ||
| The Vertebrates | Overview |
| Vertebrates Home | Vertebrate | Vertebrate |
Abbreviated Dendrogram
Synapsida
│
└─Therapsida
├─Raranimus
└─┬─Biarmosuchia
└─┬─Dinocephalia
│ ├─Anteosauria
│ └─Tapinocephalia
└─┬─Anomodontia
│ ├─Venyukovioidea
│ └─Dicynodontia
└─Theriodontia
├─Gorgonopsia
└─┬─Therocephalia
└─Cynodontia |
Contents
Index |
|
Cynognathus. Early Triassic of South Africa. From Cox et al. 1988). |
The Therapsida, the basal members of which were traditionally called "mammal-like reptiles" are the advanced synapsids, and include the mammals. The evolutionary Linnaean classification groups the therapsids into several suborders - usually Phthinosuchia/Biarmosuchia, Dinocephalia, Anomodontia, and Theriodontia, this last often subdivided. See the unit Dendrogram.
Overall, the story is as follows. Evolving from mid-Permian ancestors similar to Tetraceratops (a small synapsid completely unrelated to the well-known dinosaur Triceratops), these creatures evolved progressively more mammalian features, first in the disorderly branching of poorly known basal forms like the Biarmosuchia, Phthinosuchia, and Eotitanosuchia. From this basal group the tree developed a threefold branching. The earliest to develop were the somewhat more derived ungainly carnivores, omnivores and herbivores of the Dinocephalian lineages. Following them came two very distinct lines of adaptive evolution, the diverse and successful dicynodonts Anomodontia), and the very mammal-like theriodonts. Mammals evolved from the later group through the various intermediate stages covered here and in the next two units.
 |
Therapsida. Stratigraphic phylogeny (somewhat different from that discussed here) from Hopson (1969). |
The early members of these groups, Dinocephalia, Anomodontia, and Theriodontia are the subject of this Unit. As with so much in nature, it seems to all be about food, at least getting enough food, and processing it better. Quite simply, the three principal radiations within the Therapsida appear to be based on three solutions to the problem of bringing the food-processing surfaces of the upper and lower jaws into closer contact to achieve a more efficient system for subdividing both plant and animal foodstuffs.
The pelycosaurs (basal Synapsida) are known mainly from the Early Permian of North America (with a smattering of European forms); but this is due to accidents of preservation in the fossil record, and possibly also to restriction to the equatorial belt during the Permo-Carboniferous ice ages. Our knowledge of the therapsids is more geographically extensive. Members of all the basic therapsid lineages, apart from the Theriodonta, occur in the earliest therapsid faunas of Russia, i.e., the early Middle Permian Ocher fauna Wordian Age). It used to be thought that even more primitive fragmentary remains from the slightly earlier San Angelo formation of Texas were also therapsids Olson, 1962), but it is now known that these remains were misidentified -- these were actually caseids, not therapsids at all.
The fossil remains of the therapsids animals are also represented in the middle and upper Permian deposits of Africa, especially the spectacular Karoo formation of South Africa, from other formations in Russia, the uppermost Permian sediments of China, and from the Triassic of all of these regions. In addition, therapsids have been recovered from Europe, North America, Australia, and Antarctica. Of all the therapsids, only our own lineage, the Cynodontia, survived into the Jurassic. This group is covered in the next Unit.
MAK, slightly revised ATW051108.
As any Russian will be happy to tell you, Russia is quite different from the rest of the world; and this is especially true of the Cisuralian region. Here, waves of Indo-Europeans and Ugriks and Slavs and Tatars and Mongols and Germans and others whose very names have been forgotten have thundered like surf across the plains and forests and splintered against the base of the ancient Urals. Here, about 400 km east of Kirov, in the foothills of the Urals, is the city of Perm, which gave its name to the Permian Period. Economically, Perm has been a mining and metals town since the Sixteenth Century. Paleontologically, it is the capital of the northern Therapsida, most famous for its anteosaurs, who, like the various nations of mankind, left their remains scattered from Archangelsk to Orenburg in waves of evolutionary development during the Late Permian.
Historically, South Africa has been rather similar, with successive waves of San, !Kung, Bantu, Zulus, Dutch, English, Indians This is the capital of the southern Therapsida, who likewise left successive ecological assemblage zones in the incomparable deposits of the Karoo.
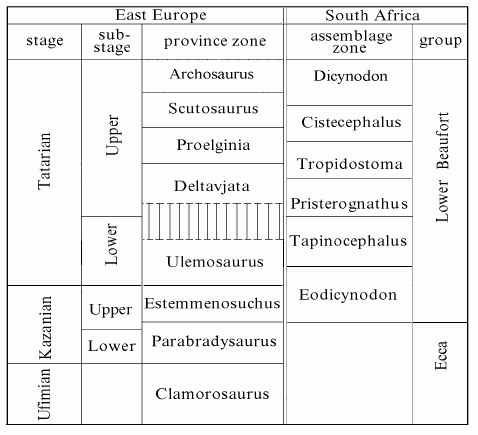
Unfortunately, Russian and South African idiosyncrasies also extend to matters of Late Permian stratigraphy. The ICS nomenclature adopts the Russian Ages for the earlier parts of the Permian, but things are more confused in later, Zechstein Time. The Russian Tatarian Age, in particular, has been synonymized with everything from the Changhsingian to the entire Zechstein. In South Africa the Lystrosaurus Assemblage Zone, starts late in in the Changhsingian and lacks the decency to stop at the Permo-Triassic boundary, as everything else in the world seems to have done. By contrast, the Capitanian, a rather bland time elsewhere, was a time of great faunal turnover in South Africa.
Accordingly, it is necessary to supplement the standard nomenclature as shown below. The ICS has not adopted dates for most of the later Permian Ages, and the literature is remarkably divergent on the equivalences between Russian, South African, and ICS time periods. Therefore, we have adopted the following, somewhat arbitrary equivalences for the terms which are emphasized in the table below. We have generally also followed the lead of Modesto & Rybczynski (2000) for Russia, and Hancox & Rubidge (1997) for South Africa.
ATW
Therapsids are difficult stuff. Evidently, some remarkably strange and interesting evolution occurred in the Late Carboniferous or Earliest Permian. At this point we have little or no idea why, but in the relatively brief period of 20 My we go from Bob the Basal Amniote (~300 Mya) to Tetraceratops ~280 Mya). We may judge how quick and uneven things were from a few basic facts of time and place. Oedalops, the most basal synapsid known from more than scraps, was contemporaneous with Tetraceratops, the most basal Therapsid known from more than scraps. Not only did they live at about the same time, but they lived within a few hundred kilometers of each other.
A glance at the main entry for Therapsida will quickly demonstrate how much had changed from even advanced pelycosaurs. For many years, in fact, the gap seemed inexplicably large. Then, in the late 1980's, Robert Reisz and his graduate student, Michel Laurin, began re-examining Tetraceratops -- a fossil which had been discovered and described as an aberrant eothyrid almost a century ago. The eventual result was Laurin & Reisz (1996). This paper is a real joy to read, like hearing two professional musicians work through a piece they know and understand. The result is smooth, compelling and complete, yet economical and understated. It is solely in an effort to avoid sounding too much like a review of Château Fombrauge, 1995 (a classic Grand Cru Bordeaux from St.-Emilion) that we will reluctantly move on from aesthetics to anatomy.
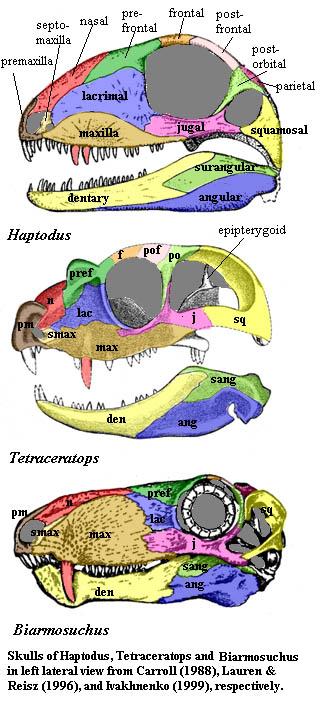
Although the skull of Tetraceratops is not quite complete, it does allow us to compare a series of fairly well known skulls:
The last requires some explanation. Ivakhnenko synonymizes Biarmosuchus tener and Eotitanosuchus olsoni. Whether or not his reconstruction corresponds to an actual creature, it at least represents a good morphotype for slightly more derived, but still primitive, therapsids from the Late Permian. Thus, these three reconstructions bracket the critical divide marking the Therapsida from other synapsids.
Marginal Dentition
We can orient ourselves with the canine teeth (in red) -- actually caniniforms, since canines are regarded as a specialized tooth form of mammals. In Haptodus the canines are not sharply set off from the other dentition. Haptodus has several precaniniform teeth on the maxilla, and the postcaniniform teeth form a graded series of decreasing size. Tetraceratops may have the therapsid condition of a single canine, although Laurin & Reisz identified a possible replacement pit for a second canine. In any event, there is only a single precaniniform tooth on the maxilla, and the canines are preceded by a marked diastema and succeeded by a series of post-canine teeth which are curved, pointed, but of much smaller size. In Biarmosuchus, there is clearly a single canine, very much larger than any other tooth. The post-canine teeth are smaller, uniform, and not markedly recurved. There are no pre-canines on the maxilla.
The premaxillary teeth show the same trend toward specialization. In Haptodus, the premaxillary teeth are unspecialized and form a graded series in size, increasing anteriorly. Tetraceratops has one large pair of premaxillary teeth, but the remainder are small and relatively uniform. Biarmosuchus has the mammal-like condition with small, uniform incisor-like teeth. Thus the basic 3-part regionalization of the marginal dentition was complete by the Middle Permian, long before the development of molariform teeth, precise occlusions, or diphyodonty.
Lower Jaw
In Haptodus, the dentary is already a very large -- perhaps the largest -- element in the lower jaw. Although the dentary does become progressively larger over the course of the Mesozoic, the Permian evolution of the therapsid condition does not seem to involve growth of the dentary so much as a progressive restriction of the angular to the posterior portion of the jaw. Note that the dentaries of Haptodus, and even Oedalops (Langston [1965]), actually extend further onto the dorsal surface of the coronoid process than do the dentaries of the therapsids Tetraceratops and Biarmosuchus. Similarly, the reflected lamina of the angular is certainly more obvious in Biarmosuchus than in Haptodus, but does not seem to be an evolutionary novelty of Therapsids.
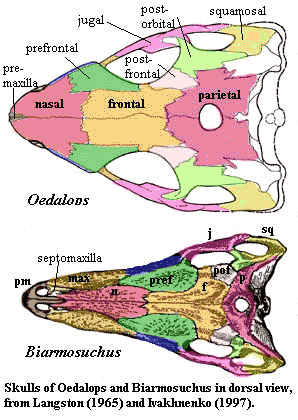
Dermal skull
Just as the dentary comes to dominate the anterior jaw, the maxilla seems to push other rostral bones to marginal positions. The lacrimal is the most obvious bone on the preorbital skull of Haptodus, extending from the orbit to the nares. In Biarmosuchus, it is reduced to a peri-orbital element, with the maxilla taking up the space formerly occupied by the anterior portion of the lacrimal.
The dorsal skull of pelycosaurs is made up of a series of blocks forming an arch, as discussed in connection with the Eothyridae. This still seems to be the basic design in Tetraceratops, and even Biarmosuchus. However, in Biarmosuchus and therapsids generally, the premaxilla is added on to the anterior end of the series, somewhat reinforced by the septomaxilla. Perhaps this is related to the development of specialized anterior dentition -- the canines and incisors.
By contrast, a great deal is going on in the posterior region, although in this case, there is perhaps less than meets the eye. The posterior skull is poorly preserved in Tetraceratops and all specimens of Biarmosuchus. However, it is possible make out the main outlines as follows:
It is instructive to compare these developments with the evolutionary changes in the diapsid clan. Diapsids generally faced less of an evolutionary bottleneck in solving the fundamental problem of the gnathostomes: how to pack in more and better muscle to drive stronger jaws. Diapsids simply had more room to maneuver, especially since most diapsid taxa have done well with flexible skulls and jaws of relatively light construction. Crocs and sphenadonts are notable exceptions. A high level of skull kinesis seems to be the rule among most other diapsids. By contrast, the synapsids seem, for whatever reason, to have been committed to a rigid skull and jaw. The therapsids solved their mechanical problem by putting an increasing amount of the muscle mass outside the dermal bones, a strategy which absolutely required a rigid skull. The transfer of muscle mass outside the skull and the related commitment to a rigid internal framework eventually had the peculiar consequence of allowing the development of larger, better supported brains.

The palate
Between Haptodus and Biarmosuchus the most obvious change is in the dentition. The marginal tooth-bearing surfaces become narrower, the marginal teeth are larger and fewer, and the palatal teeth and denticles gradually disappear. Many other changes appear to be associated with the palate becoming a more vaulted structure with a partial secondary palate. For example, the vomers fuse, broaden slightly, and develop a concave ventral surface. The palatines come to meet medially anterior to a shortened pterygoid. The interpterygoid vacuities disappear. Posteriorly, the palate seems to open up. This is likely an artifact, in part, of the increased space devoted to jaw musculature. However, the difference is so dramatic that one suspects more than that may be involved.
The quadrate and stapes have already begun shrinking at this point, long before the developments in the jaw bones which would later permit evolution of the mammalian ear. The reduction of the stapes itself is easily explained by the greater integration of the braincase with the dermal bones of the skull. This reduces the importance of the stapes in supporting the braincase ventrally. The reduction of the quadrate is more difficult to understand. One possibility is that the jaw is being supported in part by a sling of soft tissues. Certainly the miniature quadrate of later forms, such as Probainognathus (see image), seems hopelessly inadequate to the task of supporting the jaw without some such mechanism.
Moral Exegesis
It is important to separate what we see in the evolution of the therapsids from what we see of evolution within the therapsids. The range considered here -- the smallest bracket we can draw around the origin of the Therapsida -- exhibits some of the long-term trends within the Synapsida, as well some trends continued in the therapsid line. But it does not show all these trends, nor does it show them consistently.
There is, for example, no indication that the size of the temporal fenestra is increasing in any organized way. The dentary is not increasing in size. The dentary is changing shape and invading the historic anterior territory of the angular, but note that the dentary actually retreats slightly before the surangular in the passage from Tetraceratops to Biarmosuchus. Palatal dentition is generally decreasing, but the pterygoid teeth of Tetraceratops are bigger than anything that preceded them, and the palatal denticle fields seem denser than in Haptodus. The parasphenoid, likewise is proportionately larger in Biarmosuchus than in Tetraceratops.
Evolutionary trends are an excellent aid to memory. They save tremendous amounts of work in memorizing detail over thousands of species and hundreds of millions of years of time. But evolutionary trends are post hoc constructs akin to mnemonics. Like Linnean taxonomy, evolutionary trends are a good means to teach and remember -- but ultimately misleading and fundamentally flawed as explanatory tools.
Trends are not, and should never be confused with, teleology, adaptation, or even functionality. For example, within the phylogenetic range of interest, the palatines overgrow the anterior process of the pterygoid, ultimately meeting at the midline and excluding the pterygoid from surface of the anterior palate. But, repeat after me:
This does not mean that the condition of the palatines is:
- moving toward any predetermined mammalian condition,
- selected for as tending to any optimum condition, or
- adapting towards a functionally superior state.
The first statement is teleological and presumes in advance a master plan or master planner.
The second statement makes two assumptions:
The third statement is perhaps better, but presumes:
Every one of these assumptions may represent a true fact. But some of them are beyond the range of science, and the rest are not universal truths and cannot be presumed in any particular case.
Although it is speculative, the better position may be that default presumption ought to be the reverse. That is, absent evidence, we should assume that each character is more or less optimally adapted within the context of:
That sounds good, but don't we suspect that a single family of modern Texas jackrabbits released into Permian Texas would soon drive every one of these slithering sphenacodonts into phylogenetic oblivion? Maybe. Maybe not. However, that isn't the issue. We study evolution and not science fiction. No Tetraceratops could give birth to a rabbit. Evolution moves from x to x + dx. The appropriate question is whether, within the range of genetic variation available to Tetraceratops, the observed bone structures were at a local optimum for survival.
The high level answers to that question, the "why" or "why not," are probably beyond us at this point. However, it leads us in the right direction, toward issues that can be addressed. What did the palatines do? How do they interact with other elements of the palate? With diet and environment? What does this suggest about the ecology of the animal? What was likely to be the range of genetic variability? By contrast, the comparison of basal therapsids to derived therians leads nowhere. It only tempts us to invoke "trends" under the metaphysical influence of which, like a pantheon of warring godlets, evolution is supposed to take place.
ATW010824
Important Note Added in Disproof
Conrad & Sidor (2001) have recently re-reanalyzed Tetraceratops and argue that it is neither an eothyrid nor a therapsid, but rather a frazzled sphenacodont who happened to get up on the wrong side of its bedding plane. Specifically, they state that:
[d]orsal movement of the right side of the skull relative to the left suggests that the previously identified piece of skull roof is actually the zygomatic arc. Elements previously identified as opisthotic, tabular, and postorbital appear to represent the quadratojugal, squamosal, and zygomatic process of the jugal respectively. The supposed reflected lamina is a break in the angular; its orientation is the reverse of the reflected lamina in advanced sphenacodonts.
Unfortunately for the discussion above, Conrad & Sidor seem to have a very good point. We are not in a position to (yet again) analyze the mortal remains of Tetraceratops. However, as mentioned, Tetraceratops lacks several traits we associate with the therapsids.
ATW020131
Most basal, but not earliest. A number of reasonable synapsid specimens are known from the upC. However, Oedalops is phylogenetically closest to Bob. The implication is that the transition from Bob to Tetraceratops may have taken considerably less than 20 My.
Figures from Carroll 1988: 366). The original citation is Currie, PJ (1979), The osteology of haptodontine sphenacodonts (Reptilia: Pelycosauria). Paleontographica, Abt. A 163: 130-168.
This is the only point at which one might quibble with Laurin & Reisz. The septomaxilla is described as both "small" and "massive." It is said not to have a facial process and figured as having one. This is probably a stylistic, rather than substantive problem. In the nature of things, the septomaxilla is a small bone, although it is perhaps relatively bulky in Tetraceratops. The facial process in Tetraceratops does not intrude pointedly between the maxilla and nasal in the manner of later therapsids, but rather wanders aimlessly into the face and stops as if confused.
The interested or unpersuaded reader is referred to The Palatines for some counter-examples.
checked ATW031025