Anapsida: Mesosauridae
Taxa on This Page
- Mesosauridae X
The Amazing Mesosaurs

The instantly recognisable, if unlikely coloured for an aquatic animal, tiger-striped Mesosaurus, by classic paleo artist Zdenek Burian. This was one of the iconic reconstructions of prehistoric life that shaped several generations of budding paleo enthusiasts (including yours truly) MAK120322 |
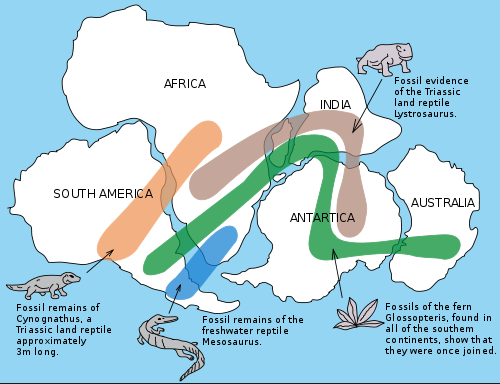
The Snider-Pellegrini Wegener fossil map, showing fossils distributed across the ancient supercontinent of Gondwana. Mesosaurus is shown at the bottom center, represented by the blue patch. This version by Osvaldocangaspadilla, Wikipedia, public domain |
Among the most distinctive of Permian reptiles are the Mesosaurs ("middle lizards"), a highly specialised group of small aquatic (or semi-aquatic) reptiles known only from the early Permian (perhaps Artinskian?) of South Africa (Namibia) and South America (Brazil, Paraguay, and Uruguay). The presence of such an endemic and stratigraphically restricted animal at opposite sides of the Atlantic ocean was a powerful argument in favour of continental drift, formulated by Alfred Wegner in the early part of the 20th century (see map at the right). Du Toit 1937
Resembling a large lizard or miniature crocodile, Mesosaurus was remarkable for its long needle like teeth, which were probably used to feed on small crustacea (they are too delicate to have been of any use for catching larger prey, such as small fish). There presence so far south during the early permian is also unusual, as much of Gondwana was still covered by glaciers. The creatures probably fed during the warm summer months, and hibernated through the winter. Their extinction would have been due to changing conditions, we could speculate that the slowly warming climate would have eventually resulted in the extinction of the specialised crustacea on which they fed
Although three genera have been named (in keeping with the tendency towards monotypal genera in vertebrate paleontology) they may be similar enough to be considered three species of the same genus; in cladistic analysis they form a closely linked monophyletic assemblage (Tsuji & Müller 2009) MAK120322
Viviparity in mesosaurs
Recently, exceptionally well preserved mesosaur embryos have been found within an adult individual, presumably a pregnant female. Several specimens also show associations between an adult and a possibly neonate. All of which suggests that mesosaurs were likely viviparous and practiced parental care (K-selection strategies). This is the earliest known case of viviparity (Piñeiro et al 2012). Such reproductive strategies have emerged independently a number of times among amniotes, as the following dendrogram shows MAK120323
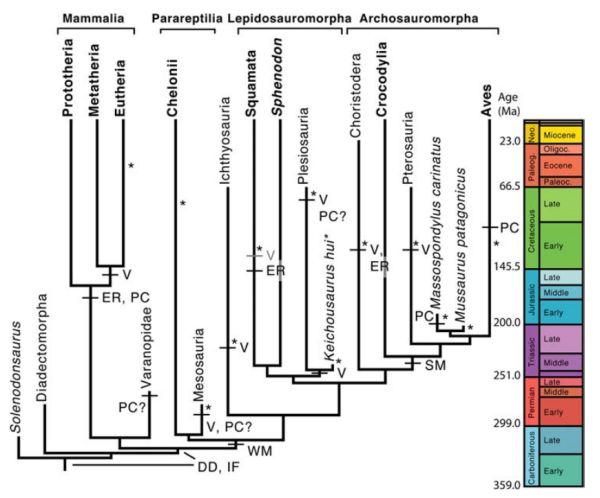
 Above: Amniote phylogeny showing the evolution of relevant developmental characters. Extant taxa in bold. Coloured timescale on right. Characters that occur only in some taxa are in grey. DD, direct development; ER, embryo retention; IF, internal fertilisation; PC, parental care; SM, strongly mineralised eggshell; V, viviparity; WM, weakly mineralised eggshell; *earliest fossilised embryos along each branch of the tree. Right, reconstruction of an adult and a posthatched Mesosaurus tenuidens, showing putative average-sized adult/neonate relation Both diagrams from Piñeiro et al 2012 Above: Amniote phylogeny showing the evolution of relevant developmental characters. Extant taxa in bold. Coloured timescale on right. Characters that occur only in some taxa are in grey. DD, direct development; ER, embryo retention; IF, internal fertilisation; PC, parental care; SM, strongly mineralised eggshell; V, viviparity; WM, weakly mineralised eggshell; *earliest fossilised embryos along each branch of the tree. Right, reconstruction of an adult and a posthatched Mesosaurus tenuidens, showing putative average-sized adult/neonate relation Both diagrams from Piñeiro et al 2012 |
Mesosaurs as diapsids? Or what?
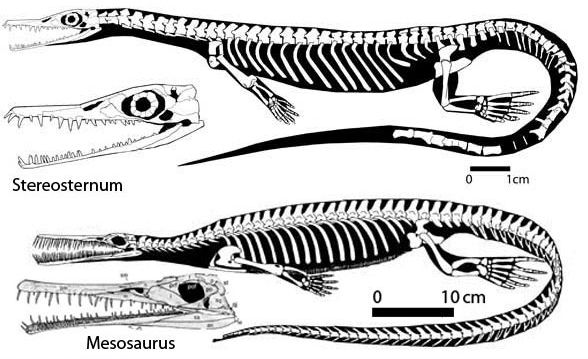
Skeletal reconstructions of the mesosaurs Stereosternum and Mesosaurus, Early Permian of Gondwana. Artwork by David Peters, used with permission |
The exact phylogenetic affinities of the mesosaurs have varied over the years. Originally they were simply included under the evolutionary linnaean Subclass Anapsida (Romer 1966, Colbert 1969, Carroll 1988), a wastebasket taxon for all reptiles that lacked temporal openings. Initial cladistic analysis showed them to be basal parareptiles (Gauthier et al 1988). Later they were reinterpreted as basal Sauropsida (as sister taxon to Reptilia) (Laurin & Reisz 1995). They are now almost universally as the most basal Anapsida/Parareptilia clade and sister taxon to all other anapsids/parareptiles (Modesto 1999b, Modesto & Anderson 2004, Tsuji & Müller 2009, Lyson et al 2010, , Tsuji et al 2012), which is basically where Gauthier et al placed them in the first place; although Hill (2005)'s total evidence tree places them even stemward of synapsids.
A basal-most taxon could mean at least one of several things:
- It really could be the most primitive or ancestral member of a group known; this doesn't seem to be the case with an animal as highly specialised as Mesosaurus.
- The basal position would be the result of a highly specialised form lack of shared apomorphies (rendering cladistic synapomorphy-based methods ineffectual. The present author (MAK) believes this is the case with placodonts in the Sauropterygia, there is also the same problem as with trying to understand Hupehsuchians), rather than a long period of cryptic evolution.
- The taxon may not belong in the group in question at all and its placement there may simply be the result of shared primitive characteristics or convergences. For example, golden moles (Superfamily Chrysochloroidea) are placed as the most basal insectivora on morphological grounds (even stemward of fossil species) but have been shown on molecular grounds to belong to a totally different group, the afrotheria.
The discovery of transitional forms would help resolve this problem. According to maverick paleo geek and artist David Peters [1], Stereosternum is less derived than Mesosaurus, and might therefore constitute such a transitional species. In email correspondence David suggests that the reason for the anomolous results in cladistic analyses would be because most researchers code Mesosaurus rather than Stereosternum, and in illustrations (right) shows the difference between the two genera., although Tsuji & Müller 2009 show all three mesosaur genera as forming a close-knit basal clade. Peters' own cladistic analysis locates the mesosaurs within the Diapsida. In view of the fact that at least one type of aeroscelid, Spinoaequalis, had become aquatic even before the Permian began, a diapsids ancestry for mesosaurids may not be unreasonable. The same result was recovered by Maisch 2010 who, like Peters, found Mesosaurus to be a sister taxon to the Ichthyosauria.However he felt this was the result of homoplasies, when he reran his analysis using a different way of coding character states, Mesosaurus was recovered in the more traditional placement of basal anapsid.
Considering mesosaurs and ichthyosaurs, two groups that are never considered together in current phylogenies, The present author is reminded of one of his childhood books on prehistoric life, it would have been circa early or mid 1960s, which had a simple evolution tree in which Seymouria was an ancestral reptile, Mesosaurus an ancestral ichthyosaur, and so on. The idea goes back to the early to mid 20th century German paleontologist Friedrich von Huene, who believed that Mesosaurus shared a common ancestry with the ichthyosaurs, deriving both from Carboniferous embolomerous amphibians, or even loxommids. Still on this subject, Huene's older American contemporary Samuel Wendell Williston placed mesosaurs, ichthyosaurs, squamates, and protorosaurs (including aeroscelids ) in a separate subclass, Parapsida, based on a shared upper temporal opening (Callaway 1997 p.5). Although the Parapsida in one form or another was still a respectable taxon in the 1950s, none of these theories are now considered to have any validity.
Although we can safely discard Huene's mesosaur-ichthyosaur association, given the huge morphological disparity and long period of time between the two groups, and the highly endemic and stratigraphically restricted range of the Mesosauridae (although Peters attempts to bridge this with several other taxa [2]), that does not mean the consensus paradigm is automatically the correct one. Given the curious topologies that regularily result through molecular phylogeny (according to which morphology is not reliable, this is a controversial subject) it is not impossible that mesosaurs may be diapsids or proto-diapsids that converged on anapsids. Of the three alternatives listed above, the present author considers the first the most unlikely, which leaves option 2 or 3. For now we have opted to follow option 2 (retained mesosaurs among the anapsids, although not as the most basal taxon), as option 3 means we have aboslutely no idea where the mesosaurs should go. MAK120323
Notes:
[1] Despite his obvious artistic talent, hard work, and dedication to unbiased research, David Peters is poorly regarded by other workers in the field. This is perhaps not so much because of his unique photo-interpretation technique, which uses Photoshop as a camera lucida, as his resulting eccentric pterosaur interpretations; see e.g. discussion on Tetrapod Zoology. In personal correspondence with the present writer (MAK) he comes across as a humble and dedicated researcher, albeit one obviously outside the mainstream. The huge cladogram on his website Reptile Evolution, shows a very deep amniota split which puts captorhinids, anapsids, diadectomorphs, turtles, and lepidosaurs in one branch, and all other diapsids, Casineria, Westlothiana, microsaurs, protorothyrids, mesosaurs, and synapsids in the other. Although parts of his tree differ from other published phylogenetic hypotheses, Peters argues that his topology is recovered when a sufficient number of taxa are included. To date no other study has attempted an analysis of the size or scope of the Peters study.
[2] Peter's results here are Stereosternum + (Wumengosaurus + (Thalattosauria + (Hupehsuchus + Utatsusaurus))). Apart from Stereosternum and the basal sauropterygian Wumengosaurus, this is a pretty uncontroversial phylogeny. Hupehsuchids are often linked with ichthyosaurs but that may be convergence. Wumengosaurus is a particularrily interesting form as its long rostrum gives it an ichthyosaur or polycotylid like appareance,
Descriptions
 Mesosauridae: Brazilosaurus, Mesosaurus, Stereosternum
Mesosauridae: Brazilosaurus, Mesosaurus, Stereosternum
Range: Early Permian of South America & Africa.
secondarily aquatic; elongated snout & neck (~10 vertebrae); nares posterior, near orbits; postorbital contacts the supratemporal; quadratojugal does not extend dorsally and is narrowly exposed on the cheek (primitive); supraoccipital lacks anterior parasagittal flanges primitive); supraoccipital plate broad & extends farther laterally than postparietal (primitive);.jaw articulation at level of occiput (primitive); ossified dorsal process of stapes (primitive); pterygoid palatal ramus extends anteriorly medial to choanae; teeth thecodont; long teeth (strained plankton?); pachostosed trunk ribs and wide neural arches, long, broad tail; well-developed anterior limbs; iliac blade with long, low posterodorsal process, lacking anterior expansion; large paddle-foot pes.
Image: Stereosternum from Permian Tetrapods. We recommend a visit to this site which has some excellent images in which some of the listed characters can be seen.
Links: Phylogeny and Classification of Amniotes; MESOSauria; Nathis Fauna Reptilelen (Dutch). ATW060129.
checked ATW040118, revised MAK120323
