Aves: Ornithothoraces
Taxa on This Page
- Carinatae
- Enantiornithes X
- Hesperornithiformes X
- Ichthyornithiformes X
- Ornithothoraces
- Ornithurae
- Patagopterygiformes X
Half-way to Herons
 The ornithothoracines are a half-way station between dromaeosaurs and neornithines -- a good place to pause and reflect on one or two points about evolution, including what it means to be "half-way" to anything. The orthodox answer is that it means nothing at all. In fact, the whole idea of being half-evolved is a dangerous and highly misleading concept. Generally speaking, this rejection is a true saying and worthy to be heard by all men (a phrase from an orthodoxy which itself is now obsolete and regarded as error).
The ornithothoracines are a half-way station between dromaeosaurs and neornithines -- a good place to pause and reflect on one or two points about evolution, including what it means to be "half-way" to anything. The orthodox answer is that it means nothing at all. In fact, the whole idea of being half-evolved is a dangerous and highly misleading concept. Generally speaking, this rejection is a true saying and worthy to be heard by all men (a phrase from an orthodoxy which itself is now obsolete and regarded as error).
Half-wayness, in fact the idea that there is any evolutionary trajectory to be half-way on, leads to a number of false concepts. It suggests teleology: that is, that a taxon is predestined to reach some particular goal. The nature of the assumed teleological force is often theological or spiritual. There is nothing necessarily false or anti-scientific about such beliefs. However, they are decidedly non-scientific. They are outside the scope of scientific investigation, which seeks to determine relationships among observable, testable phenomena. Science does not require classical causation to be valid. It does, however, assume that relationships among observable phenomena can be delimited and described in at least a statistical way and that the descriptions can be tested precisely because they have testable limits. These assumptions may or may not be true (another slippery concept) but anything that violates the assumptions is not science.
A second difficulty with the notion of trajectory in evolution is more subtle. In the discussion of alvarezsaurids, we reviewed the very rapid evolution of isolated island birds which lose the ability to fly. In this circumstance, there is in fact a clear evolutionary trajectory. Once flight is lost, there is a strong selective pressure to make certain modifications, some of which are statistically quite predictable. It is reasonably certain that the first bird to lose flight in such cases is less well-adapted to its environment than its descendants. It is, in some meaningful way, "half-evolved." However, this is a very different case from the usual discussion of broad evolutionary movements across millions or tens of millions of years. Movement across such lengths of time is very unlikely to result from some "correction" of significant maladaptation. The island bird exception proves the rule. Adaptation happens quite quickly, even after a severe population bottleneck restricts the initially available genetic variation. (How many finches originally reached the Galapagos? In all likelihood, the number was exactly two, and certainly not much higher.)
Across millions of years, the better assumption is that the species is very well adapted to its environment at all times. Thus there can be no evolutionary trajectory. Rather, we should look first to long-term changes in the environment, rather than maladaptation, to explain long-term changes in a taxon. The idea of evolutionary trajectory, of a "half-evolved" taxon, points us in the wrong direction. The longer the period under consideration, the surer we can be that the taxon was fully evolved at all times and better adapted to its environment and niche than it would have been had it possessed the new and improved version of whatever structures are under consideration. When we consider the 80-90 My between Archaeopteryx and the end-K event(s), it makes no sense to presume that Iberomesornis or any early ornithothoracine was simply a half-evolved neornithine. Rather, the logical focus is to determine what it was about the late Mesozoic environment, biotic and otherwise, that these birds were well-adapted to. Neornithines probably existed world-wide from the early Cretaceous. Had more basal ornithothoracines been significantly less well-adapted, less "fit", they would undoubtedly have been eliminated in no time at all, geologically speaking.
Another "force" which can mimic long-term evolutionary trajectories is a combination of asymmetric probabilities. The simplest case is the loss of a structure. Many genetic accidents can result in the loss of a toe. Polydactyly -- the addition of a toe -- is rarer. Given completely random genetic events, genetic drift ensures that any given taxon will lose toes over long periods of time. In cases such as this, selection can stabilize the number of digits. Three or five toes may be optimal for balance or mobility, for example, so that two- or four-toed variants are selected out of the population. But if selection is absent, the drift towards fewer toes will continue. It is the absence of selection which allows a long-term genetic change.
Postscript: This theme is further elaborated in a very different context, at Therapsida
Opposite Birds
One of the more interesting features of the enantiornithines, from an evolutionary perspective, is that they co-existed with relatively modern birds for approximately 70 My. Early ornithurine and enantiornithine birds are both found shortly after Archaeopteryx, in the earliest Cretaceous. In some respects, the evolution of the enantiornithines parallels that of the ornithurines. Over time, both groups show a gradual loss of teeth and of wing claws. The flight apparatus becomes more robust, and both develop a true fused carpometacarpus.
However, some significant differences remained stable. For example, no enantiornthine has ever been found with uncinate processes. Uncinate processes are small projections from the ribs which, in living birds, are involved in the attachment of muscles responsible for the unique avian respiratory system. While this is a long story in itself, the important point is that the lungs of birds have no elastic properties. Mammals largely rely on the elasticity of the lung to expel air. Birds (and all archosaurs, so far as it is known), must force air out of the lung. Modern birds depress the sternum which, in turn, forces air out of the abdominal air sacs and into lungs. The absence of uncinate processes suggests that the pattern of respiration may have differed materially between enantiornithine and ornithurine birds. The relatively small keel on the sternum of enantiornithines may confirm this suggestion. While such indications can hardly be regarded as proof, it is tempting to speculate that the enantiornithines either had a somewhat different method for powering respiration, or simply required less oxygen. In fact it has been suggested, for these reasons among others, that enantiornithines may not have been fully endothermic. One other indicator, the "growth" patterns in bone microstructure, has a somewhat checkered record. To the extent it remains a credible marker, this factor also indicates a more ectothermic metabolism.
Another interesting difference is one of degree. Several writers have remarked on the fact that enantiornithines had a strong reversed hallux and large, sharp claws indicating an ability (and presumably tendency) to live in trees "like modern birds." In fact it could fairly be argued that, by and large, enantiornithines had feet which are actually stronger, more flexible, and more strongly clawed than the feet of contemporary ornithurines. It may be that enantiornithines were not simply able to live arboreally, but actually dominated that environment and out-competed their neornithine cousins in this niche.
The Hesperornithiformes
 The only published fossil evidence for the presence of this group in the early Cretaceous are the somewhat ambiguous remains known as Enaliornis. However, it is possible, in fact likely, that the Hesperornithiformes are a fairly ancient group. They are clearly not Enantiornithines, as evidenced by the distal to proximal pattern of fusion in the leg bones. Thus their basic stock diverged from the more common birds of the Lower Cretaceous fairly early on. Although Hesperornis is flightless, its remains have been found in Antarctica, South America, the United States and Northern Canada. Other Hesperornithiforms (and possibly Hesperornis) have been found in Eurasian sites, including Mongolia, Kazakhstan and Sweden.
The only published fossil evidence for the presence of this group in the early Cretaceous are the somewhat ambiguous remains known as Enaliornis. However, it is possible, in fact likely, that the Hesperornithiformes are a fairly ancient group. They are clearly not Enantiornithines, as evidenced by the distal to proximal pattern of fusion in the leg bones. Thus their basic stock diverged from the more common birds of the Lower Cretaceous fairly early on. Although Hesperornis is flightless, its remains have been found in Antarctica, South America, the United States and Northern Canada. Other Hesperornithiforms (and possibly Hesperornis) have been found in Eurasian sites, including Mongolia, Kazakhstan and Sweden.
The legs of Hesperornis are extraordinarily derived. The femur is quite short and attaches to the pelvis rather far back on the long body. Since it articulates at two points with the pelvis, the femur is constrained to stick more or less straight out laterally and at right angles to the body when flexed. While the femur could likely fold back against the side to a considerable degree, it cannot rotate dorsoventrally. Thus, the feet also faced outward, with the long, broadly lobed toes pointing sideways. In this posture -- similar to a penguin, but more extreme -- it is unlikely that the bird could walk at all. This has led to considerable speculation about just how and where it laid eggs. The current legend is that Hesperornis crawled onto select, isolated islands which it used as rookeries. However, actual evidence is slim to non-existent. As in mosasaurs, the mechanics of egg-laying (if any) and prenatal care (if any) are not easy to imagine.
The diving movement of Hesperornis is not entirely clear. The cnemial crest is a rather enormous proximal extension of the tibiotarsus and sticks out beyond the distal end of the femur. This feature is convergent with modern diving birds and probably indicates that the musculature attaching to the cnemial crest was sufficiently massive that, given the short femur, the upper leg was virtually incorporated into the body wall. In modern divers, such as the penguin and loon, the wings are also shaped so that the surface presented to the water is a smooth, hydrodynamic curve. In Hesperornis, the wings are vestigial, and the femur relatively shorter and more posterior, giving the bird, in life, perhaps a the appearance of an acute triangle (reconstructions showing a large-breasted teardrop form are common, but may be mistaken). During the recovery stroke, the femur must have been drawn forward, the muscles attaching to the tibiotarsus relaxed, and the toes rotated so that the blade of the lobes cut almost frictionlessly through the water. As a natural result, the lower leg would be drawn closer to the body. At the end of the recovery stroke, the toes are rotated flat to the water, like an oar, and the bird "steps" outward with both legs, with a resultant force that shoots it forward.
If this is correct, then lateral diving control would be a simple matter of adjusting the relative strength of the kick in each leg. Lacking wings, however, it is less easy to see how attitudinal control was achieved or how the bird could adjust direction without accelerating. Turning is more economically achieved by small forces distant from the center of gravity, such as those exerted by wings. Compared to modern divers, then, the hesperornithiforms may have been sprinters, rather than long distance pursuit hunters; able to accelerate rapidly and use their slender bodies and long necks to snatch prey, rather than running it down and outmaneuvering it. ATW000130
Ornithothoraces: Iberomesornis, Otogornis, Platanavis. Definition: Iberomesornis + Neornithes. Padian et al. 1999), [C95]
uncinate processes absent primitively; gastralia absent [P+93]; pelvic bones fused ?), but individual 3-4 sacral vertebrae in early forms, rather than larger, more closely fused synsacrum; $ pygostyle present [C95]; primitively, pygostyle from fusion of distal caudals at end of short tail; $ coracoid strap-like [P+93] [C95]; $ coracoid forms sharp angle with scapula at level of glenoid [P+93]; $ scapula tapers gradually [P+93]; $ scapula terminates distally in sharp point [C95]; well-developed U-shaped furcula [P+93] with hypocleideum in flying forms; $ humerus shorter than ulna [P+93]; $ shaft of ulna twice as thick as radius [P+93] [C95]; distal carpals may be unfused; manus generally modern; tarsometatarsus largely unfused, esp. distally, in early forms; reversed hallux; all with essentially modern feathers.
Links: Aves; Avialae -- The Dinosauricon; Paleontology and Geology Glossary: I.
References: Chiappe (1995) [C95]; Perle et al. 1993) [P+93]
The diverse and successful "opposite" birds, so called because the tarsometatarsus is fused proximal to distal and because the triosseal canal is formed primarily by the scapula (rather than coracoid). teeth primitively present; uncinate processes absent; coracoid strut-like, as in Neornithines; sternum keeled, but keel may be restricted to posterior of sternum; elongate hypocleideum; fused carpometacarpus; at least 8 vertebrae in synsacrum; hind limbs diverse and often primitive; true tibiotarsus, tarsometatarsus fused only proximally; tarsal cap absent; fully reversed hallux with large hallical claw; often strongly curved, sharp claws. Pattern of bone growth has suggested to some that * may have been, if not ectothermic, at least less than fully endothermic. Embryos probably precocial.
Links: Subclass Enantiornithes; Dann's Dinosaur Info; Aves Tree of Life Chiappe article); Aves Translation and Pronunciation Guide; New Scientist Planet Science | The Rex Files; EARLY BIRD FOSSIL; Enantiornithes -- The Dinosauricon.
 Ornithurae:
Apsaravis, Gansus, Chaoyangia, Ambiortus, Liaoningornis. Definition: Hesperornis + Neornithes. Padian et al. (1999).
Ornithurae:
Apsaravis, Gansus, Chaoyangia, Ambiortus, Liaoningornis. Definition: Hesperornis + Neornithes. Padian et al. (1999).
Range: from the Early Cretaceous
Relatively modern birds. $ quadrate with sharp orbital process [C95]; heterocoelous cervicals [ZZ01]; $ less than 11 dorsal vertebrae [C95]; ribs with uncinate processes; synsacrum composed of 9 or more sacrals [ZZ01]; pygostyle short [ZZ01]; scapula curved [ZZ01]; triosseal canal formed largely from coracoid; coracoid with a well-developed procoracoid and a round scapular articulating fossa [ZZ01]; paired arms of furcula more parallel & presumably capable of being compressed as spring-like energy storage device; sternum elongate with antero-lateral processes, keel high and extending along the full length of the sternum [ZZ01]; broad sternum presumably capable of powering air sac system; keel approaches anterior margin of sternum; ulna with a semicircular dorsal condyle [ZZ01]; two distal metacarpals fused; first phalanx of second manual digit antero-posterioly expanded [ZZ01]; $ small acetabulum [C95]; $ pubis & ischium more or less parallel and directed posteriorly [C95]; pubes separated distally; small acetabulum; femur with prominent patellar groove [C95]; tarsal "cap," distal to proximal pattern of fusion in tarsometatarsus; tarsometatarsus completely fused [ZZ01]; bone usually fibrolamellar and lacks "growth" rings; Mesozoic forms reportedly found in marine locations far from nearest land.
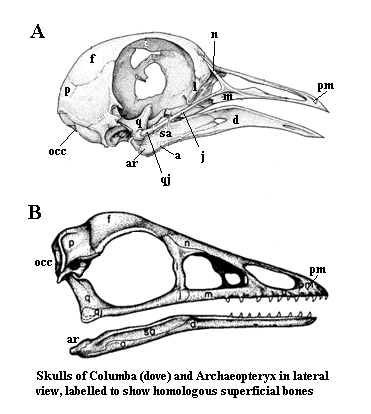
.Image: Right: Columba modified from Procter & Lynch (1993). Archaeopteryx modified from Feduccia 1999) (citing Martin, LD (1991), Mesozoic birds and the origin of birds in H-P Schultze & L. Treube (eds.), Origins of the Higher Groups of Tetrapods, Cornell Univ. Press: pp. 485-540.) a angular; ar articular; d dentary; f frontal; j jugal; l lacrimal; m maxilla; n nasal; occ occiput; p parietal; pm premaxilla; q quadrate; qj quadratojugal; sa surangular. Left: Yanornis from [ZZ01].
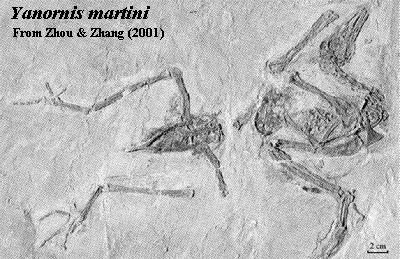 Note: In addition to the reorganization of the jaw, note how the entire top of the skull has been rotated and posteriorly expanded. In the pigeon Columbia), the upper jaw is movable on the quadrate and flexes on the thin bones of the jugal and maxilla, and at a joint at the frontal-nasal articulation. This appears to be a synapomorphy of the Ornithurae, although other groups may have developed different modes of kinesis.
Note: In addition to the reorganization of the jaw, note how the entire top of the skull has been rotated and posteriorly expanded. In the pigeon Columbia), the upper jaw is movable on the quadrate and flexes on the thin bones of the jugal and maxilla, and at a joint at the frontal-nasal articulation. This appears to be a synapomorphy of the Ornithurae, although other groups may have developed different modes of kinesis.
Patagopterygiformes: Gargantuavis, Kuszholia, Patagopteryx.
Range: Early(?) Cretaceous of South America, possibly Asia.
Characters: chicken-sized; quadrate with 3 distal condyles forming a triangle; unique fused quadrate-pterygoid complex; 3rd pair of transverse processes enlarged; synsacrum present, wide & ventrally convex; caudal prezygapophyses absent or much reduced; very small wings (flightless) distinct neck posteroventral to the glenoid; absence of pubic foot; tarsometatarsal vascular distal foramen.
Links: Patagopteryx
-- The Dinosauricon; Patagopteryx; Untitled; Aves Translation and Pronunciation Guide; SAPE Newsletter 1993 no.7; Senckenberg Peters- Die Entstehung der Vögel; Patagopteryx. ATW040306.
 Hesperornithiformes:Enaliornis?, Hesperornis.
Hesperornithiformes:Enaliornis?, Hesperornis.
Loon-like aquatic birds up to 2 m, but notably long and thin, incapable of flight and probably incapable of walking. Nasal turbinates present; teeth absent from premaxilla, but present on maxilla and dentary; keratinous beak present in some or all species; beak may have been present on upper jaw only (H.) (see All About Archaeopteryx); skull generally as in modern birds, but with large vomer, possibly intramandibular joint with predentary; supraorbital salt gland may have been present; vertebrae amphicoelous; anterior ribs not attached to sternum; sacrum long; tail short; sternum lacks keel; wings vestigial, pelvis very long and narrow; pubis and ischium fully retroverted and parallel to sacrum; ischium not fused to ilium; femur oriented laterally, flexed tibiotarsus oriented caudally; femur has accessory articulation to pelvis represented by indentation next to acetabulum; femur very short; tibiotarsus long, with long cnemial crest; laterally compressed tarsometatarsus; pattern of fusion in legs is distal to proximal, feet large; individual digits lobate, not webbed; toes able to rotate (thus reducing drag on recovery stroke); unclear whether LAGs "growth" rings) present in bone; bones not pneumatic, although air sinuses present in cranial bones; feathers were plumaceous and hair-like in Parahesperornis.
Links: Hesperornis -- The Dinosauricon; Hesperonis; Image -- The Dinosauricon; cretaceous images page 2; Lectures on Evolution (1877) (Lecture by "Darwin's Bulldog"); Other Replicas.
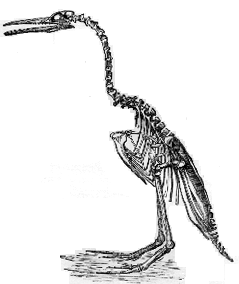
Image: The image is Hesperornis regalis from a monograph by OC Marsh. It suggests a posture that the bird could probably not have assumed in life. Current reconstructions might also place the sternum somewhat more posteriorly. [Possibly Marsh, OC (1880) Odontornithes: a monograph on the extinct toothed birds of North America, Professional Paper, Engineering Department, United States Army -- although the drawing itself is apparently older and may derive from an 1872 paper I have been unable to locate].
 Carinatae: Definition: Ichthyornis + Neornithes. Padian et al. (1999), [C95]. Traditionally, birds with keeled (i.e. a prominent medial, longitudinal ridge) sterna.
Carinatae: Definition: Ichthyornis + Neornithes. Padian et al. (1999), [C95]. Traditionally, birds with keeled (i.e. a prominent medial, longitudinal ridge) sterna.
Range: from the Early(?) Cretaceous
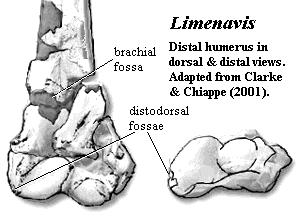
Characters: $ globe-shaped, cranio-caudally convex humeral head [C95] [CC01]; $ brachial fossa of humerus [CC01]; $ one or two distodorsal humeral fossae [CC01]; $ complete proximal and distal metacarpal fusion [CC01]; $ extensor process on metacarpal I [CC01]; $ extensor groove on distal tibiotarsus [CC01].
Note: Traditionally, "Carinatae" was used to mean all neornithines other than ratites. This is more or less equivalent to Neognathae in many cladistic schemes (that clade is unnamed in these Notes).
Links: Aves/_MORPHOLOGIE/skelett01.html">Bestimmungsuebungen an Wirbeltieren (German); Ornithurae -- The Dinosauricon; Lectures 16-18: Early Cretaceous.
References: Chiappe (1995) [C95], Clarke & Chiappe (2001) [CC01]. ATW031010.
 Ichthyornithiformes:
Ichthyornis.
Ichthyornithiformes:
Ichthyornis.
Stout, gull-like shore birds with large heads and wings, up to 0.5 m. Supraorbital salt gland may have been present; intramandibular joint as in Hesperornithiformes; teeth apparently in open groove & sockets developed ontogenetically; sternum well-developed and carinate; wings large and muscular; feet small.
Links: Ichthyornis -- The Dinosauricon; Ichthyornis Printout- EnchantedLearning.com; Other Replicas; Lectures on Evolution (1877); National Geographic Magazine: July 1996 @ nationalgeographic.com; Dinosaurs and Birds - Enchanted Learning Software.
 The ornithothoracines are a half-way station between dromaeosaurs and neornithines -- a good place to pause and reflect on one or two points about evolution, including what it means to be "half-way" to anything. The orthodox answer is that it means nothing at all. In fact, the whole idea of being half-evolved is a dangerous and highly misleading concept. Generally speaking, this rejection is a true saying and worthy to be heard by all men (a phrase from an orthodoxy which itself is now obsolete and regarded as error).
The ornithothoracines are a half-way station between dromaeosaurs and neornithines -- a good place to pause and reflect on one or two points about evolution, including what it means to be "half-way" to anything. The orthodox answer is that it means nothing at all. In fact, the whole idea of being half-evolved is a dangerous and highly misleading concept. Generally speaking, this rejection is a true saying and worthy to be heard by all men (a phrase from an orthodoxy which itself is now obsolete and regarded as error).