
Life in the Devonian fast lane: Dunkleosteus telleri prepares to seize a Gorgonichthys clarki, while a Coccosteus cuspidatus, and a school of Ctenurella gladbachensis swim by. A trio of Strunius walteri coelacanths flee. © Stanton F. Fink
| Placodermi | ||
| The Vertebrates | Overview |
| Vertebrates Home | Vertebrate | Vertebrate |
|
Abbreviated Dendrogram
Gnathostomata
│ └─Acanthodii etc (if traditional placement)
│
└─Placodermi
╞?═Acanthothoraci
├─Rhenanida
└─┬─Antiarchi
│ ├─Yunnanolepidoidei
│ └─EuantiarchaAntiarchi
└─┬─┬─Petalichthyida
│ └─Ptyctodontida
└─┬─┬─Arthrodira
? ├─Actinolepidoidei
│ │ └─Phyllolepida
│ └─┬─Phlyctaenioidei
│ └─┬─Brachythoraci
│ └─┬─Coccosteomorphi
│ └─┬─Pachyosteomorphi
│ └─Aspinothoracidi
╘═Acanthodii
├─Chondrichthyes
└─Osteichthyes
|
Contents
Overview |
|
Life in the Devonian fast lane: Dunkleosteus telleri prepares to seize a Gorgonichthys clarki, while a Coccosteus cuspidatus, and a school of Ctenurella gladbachensis swim by. A trio of Strunius walteri coelacanths flee. © Stanton F. Fink |
 |
| Bothriolepis. Late Devonian - worldwide. Length about 20 to 30 cm |
The name "placoderm" is from the Greek and means "tablet + skin", referring to the heavy armoured bony plates that completely covered the head and thorax of these curious prehistoric fish. These creatures are among the most ancient of jawed fish, and were previously thought to be, along with the Acanthodii, the only class of (gnathostomes) to become completely extinct. A recent study however (Brazeau 2009) suggests that neither of these groups became extinct, rather, they are paraphyletic/ancestral groups that evolved into higher fish. This is less of a radical suggestion with the acanthodii or "spiny sharks", long suspected to being a paraphyletic (ancestral) group. It is more controversial regarding the placoderms, who were much more specialised, with many unique features (in cladistic jargon synapomorphies) not shared by other vertebrates.
With their armored head shield and trunk shield composed of overlapping bony plates, the placoderms appear at first glance extremely similar to the ostracoderms (armoured jawless fish). They are easily distinguished however by their paired fins and presence of jaws, an adaptation that made them quite different from their earlier jawless cousins, for it enabled them to bite solid food rather than simply suck up organic particles from the mud. Yet still these creatures were equally different from othe jawed fishes. The jaws were supported in a distinct way, with the palatoquadrate (upper jaw bone) fitting inside the arms of the lower jaw. This gave the placoderms a distinctively narrow gape. However, the head and trunk shields of most placoderms were articulated by bony joints, allowing the forward part of the skull to tilt up, increasing the gape and allowing them to take in larger food. It used to be believed that placoderms lacked teeth and relied on biting or grinding structures formed by dermal bones lining their mouths. The current view is that these are true teeth, formed by fusion of multiple dental elements as in extant holocephalians. The backbone consisted of a notochord (an embryonic feature in higher vertebrates) that persisted throughout life, and vertebra consisting only of Y-shaped spines above and below it. These were often cartilaginous, like the skeleton of sharks (Chondrichthyes), a group with which the Placoderms have sometimes been considered to share a common ancestry.
Although the head and thorax were usually armored with dermal bone, the rest of the body was quite vulnerable, covered with small bony scales or lacking even that. It is not clear what purpose this armour may have served. It has been suggested that it was a defense against the giant sea-scorpions that inhabited brackish water environments around this time. Another explanation is that it may have served as an exoskeleton and (as with arthropods and mollusks) a support for the external organs (in view of the very weak cartilaginous skeleton this may not be unlikely)
Most placoderms were quite small, about 10 or 15 cm in length, but one important clade, the Brachythoraci, included both carnivorous and filter-feeding types types and reached lengths of upto 4 to 9 meters in length, making them the largest animals of the Devonian. Images of the enormous carnivore Dunkleosteus, a representative type, are common on the web.
Although they first appeared during the early Silurian period, placoderms did not become common or widespread until the Early Devonian, when they underwent an extraordinary evolutionary radiation. They soon came to dominate most brackish and near-shore ecosystems, and also spread to marine and freshwater environments. More than 250 genera are known in all, with almost 100 species of one genus, Bothriolepis, making them the most diverse and important of Devonian vertebrates.
The placoderms were seemingly little affected by the extinction at the end of the Frasnian Age, yet the group dwindled during the Famennian and died out at the Devonian/Carboniferous boundary.
Text by MAK. Revised ATW050825, MAK111028.
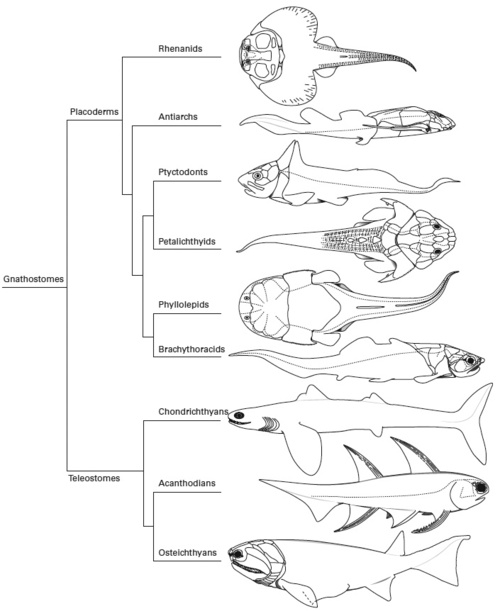 Diversity of Placoderms. Reconstructions of representative taxa placed on a dendrogram. Note that placoderm phylogeny are still disputed, see dendrograms page for some different interpretations. From top to bottom: Gemuendina stuertzi, a rhenanid, in dorsal view; Bothriolepis canadensis, an antiarch, in lateral view; Ctenurella gladbachensis, a ptyctodont, in lateral view; Lunaspis broilii, a petalichthyid, in dorsal view; Phyllolepis orvini, a phyllolepid arthrodire, in dorsal view; and Coccosteus cuspidatus, a brachythoracid arthrodire, in lateral view. Also shown are three representative non-placoderms; Cladoselache fyleri, a chondrichthyan, in lateral view; Diplacanthus ellsi, an acanthodian, in lateral view; and Cheirolepis canadensis, a paleoniscid osteichthyan, in lateral view. From Scientific Illustration blog, diagram originally from The vertebrate fauna of the Cleveland Shale (Famennian, northern Ohio), Ohio Geological Survey, Chapter 5, Field Guide 22. by Carr, R. K. and G. L. Jackson |
 Lifestyles of the Peculiar and Armor-plated: In a lake in Devonian Gondwana, an Austrophyllolepis ritchiei settles in while a trio of Bothriolepis askinae forage, and a school of Culmacanthus antarctica swim by. © Stanton F. Fink Stanton 060924 |
Placoderms evolved into a variety of ecomorphs (body types), including torpedo-shaped swimmers, flattened bottom-dwellers, and armoured box-like types. They included both carnivorous and detritivorous types. The pared fins and a heterocercal tail suggest that they could swim quite efficiently when they wanted to, but the heavy armour would have weighed them down, and it is likely they spent much of their time sitting on the bottom.
Although the head and thorax were heavily armored with dermal (skin) bone, the rest of their bodies were quite vulnerable, covered with small bony scales or lacking even that. It is not clear what purpose their armour may have served. It has been suggested that it was a defense against the giant sea-scorpions that inhabited brackish water environments around this time. Another explanation is that it may have served as an exoskeleton and (as with arthropods and molluscs) a support for the internal organs (in view of the very weak cartilaginous skeleton this may not be unlikely)
Many placoderms, particularly the Rhenanida, Petalichthyida, Phyllolepida, and Antiarchi, were bottom-dwellers. As such, Placodermi has been popularly misinterpreted as being a tribe of bottom-feeding snails and garbage trucks, nevermind that the placoderms were the dominant vertebrate group during the Devonian. One must remember that the vast majority of placoderms were predators, many of which lived at or near the bottom. Many, primarily the Arthrodira were mid- to upperwater dwellers, and were active predators. The largest known arthrodire, Dunkleosteus telleri, was a 6 to 8 meter long predator and was presumed to have a nearly worldwide distribution, as its remains have been found in Europe, North America and Morocco. Other, smaller arthrodires, such as Fallacosteus and Rolfosteus of Gogo, had streamlined, bullet-shaped head armor, strongly crediting the idea that many, if not most, arthrodires were active swimmers, rather than passive ambush-hunters whose armor practically anchored them to the seafloor.
It was originally thought that the placoderms went extinct due to competition from the first bony fish, as well as the early sharks, due to a combination of the supposed inherent superiority of the bony fish and sharks, as well as the presumed sluggishness of the placoderms themselves. Since then, though, as more accurate summaries of prehistoric organisms have been developed, it is now presumed that the last placoderms died out one by one as each of their ecological communities suffered due to the environmental catastrophes during the Devonian/Carboniferous extinction event. MAK060926
The earliest studies of placoderms were published by Louis Agassiz, in his five volumes on fossil fishes, 1833 – 1843. In those days, the placoderms were thought to be shelled jawless fish akin to ostracoderms. Some naturalists even suggested that they were shelled invertebrates, or even turtle-like vertebrates. The work of Dr. Erik Stensio, at the Swedish Museum of Natural History, Stockholm, from the late 1920s established the details of placoderm anatomy, and identified them as true jawed fishes related to sharks. Using the Sollas method, he took fossil specimens with well-preserved skulls, and ground them away, one-tenth of a millimeter at a time. Between each grinding, he made an imprint in wax. Once the specimens had been completely ground away (and ironically, completely destroyed as a result), he made enlarged, three-dimensional models of the skulls in order to examine the anatomical details more thoroughly. Many other placoderm specialists suspected that Stensio was trying to shoehorn placoderms into a relationship with sharks, but with with the discovery of more fossil specimens, especially the exceptionally well-preserved fossils from the Gogo Reef formation in Australia, Stensio's theory of sharks and placoderms as sister groups is accepted as fact due to a whole host of similarities between the skull anatomies of the two groups.
The first identifiable placoderm fossils are known from the Middle Silurian (Wenlock). The first appearance of Silurian placoderm fossils, in China, show the fishes already differentiated into Antiarchs and Arthrodires, along with the other, more primitive groups; apparently Placoderm diversity originated long before the Devonian, somewhere in the middle Silurian, though earlier fossils of basal Placodermi, have yet to be discovered in these particular strata. At least, that's what scientists assume so far. The Silurian placoderm fossil record is literally fragmented, as they are known only from fragments of plates and armor. Some of these fragments bear a strong enough resemblence (due to ornamental and or histological details) to certain groups (i.e., the Antiarchs and Arthrodires) to be identified as being members of those groups, though none of these fragments have been officially described, or even named. Ironically, the best known, or at least, most commonly cited example of a Silurian placoderm, Wangolepis, is known from fragments that currently defy attempts at placement in any of the known orders of placoderms.
MAK060926, MAK060930
The UCMP site has coverage equivalent to ours -- better organized but now a bit dated. Steven Carr's placoderm page has the placoderms as paraphyletic ancestors of the sharks. We're not sure where this comes from. The internal arrangement of the various placoderm groups is unclear. Compare Carr's phylogeny with the two trees in Mikko's Phylogeny. ATW
 |
| Bothriolepis canadensis © University of Oslo Paleontological Museum. Reproduced by permission. |
lwS-upD.
Gnathostoma: Eugnathostoma + *: Arthrodira + (Petalichthyida + Ptyctodontida) + (Rhenanida + Antiarcha).
Teeth dermal bone, not replaced; jaw muscles medial to palatoquadrate; nostrils anteroventral; nasal capsules not fused to braincase; stalked eyeballs (as chondrichthians); sclerotic plates of 1-4 pieces; spiracle absent; probably 5 branchial arches; single branchial opening between cephalic & thoracic armor; unique neck joint; head and (variably) body covered with dermal bone plates; pattern of plates characteristic of each placoderm order; neural arches present and, in the tail region, hemal arches (no ossified centra); heterocercal tail; single dorsal fin; anal fin unknown or absent; internal skeleton of cartilage and perichondral bone; well-developed and unique pectoral fins; primitively, presence of semi-dentine with tear-drop shaped cavities for encased odontocytes; body often flattened (benthic).
Introduction to the Placodermi Extinct armored fishes with jaws - a good non-technical coverage; "Devonian Armoured Fish" Annetta Markussen-Brown, 2000; Palaeozoic fossils UK; Placodermi - armoured sharks, University of Oslo, Norway; Animal Diversity Web: Placodermi phylogeny; Dunkleosteus sp. Devonian Placodermi from Morocco; Bothriolepis, the most successful of all the placoderms with some 100 species known from every continent; Mikko's Phylogeny Archive - Placodermi - dendrograms; Coccosteus site - Placodermi (Italian); Placodermi - the only entirely extinct class of vertebrates; Wikipedia - Placodermi - some of this material was incorporated into these pages, and vice versa. checked HAJ090203.
Note: The body and head armor are primitively not articulated, with several independent lines developing articulated armor. ATW010420
checked ATW0151003, revised MAK111028