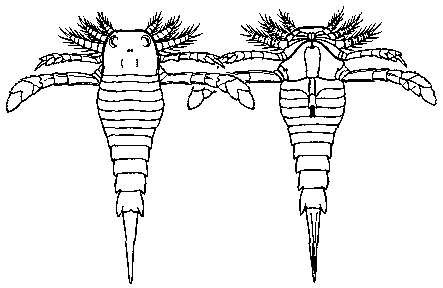
Dolichopterus macrocheirus
length 25 to 30 cm
Late Silurian period of west central Euramerica
| Eurypterida | ||
| Arthropoda | Eurypterida |
| Metazoa | Metazoa | |||||
| Glossary | Time |
|
Chelicerata
├─┬─Xiphosura
│ └─Eurypterida
│ ├─Eurypteracea
│ │ ├─Mixopteracea
│ │ └─Pterygotina
│ └─Stylonuracea
│ ├─Dolichopteridae
│ └─┬─Stylonuridae
│ └─Hibbertopteroidea
└─Arachnida
├─Scorpioniformes
│ ├─Opiliones
│ └─Scorpionida
│ ├─Palaeoscorpiones
│ └─Orthosterni
│ ├─Chactoidea
│ └─Scorpionoidea
└─Araniformes
|
|
Dolichopterus macrocheirus
length 25 to 30 cm
Late Silurian period of west central Euramerica
The Eurypterida are an extinct Paleozoic group of chelicerate arthropods of which 200 fossil species are known. These were spectacular animals, although very rare as fossils. The largest, such as Pterygotus; reached 2 meters and more in length, but most species were less than 20 cm. Although called "sea scorpions" only the earliest ones were marine. Most lived in brackish water, sheltered lagoons etc. Many species inhabited shallow aquatic environments and some may have been amphibious, emerging onto land for at least part of their life cycle. They may have been capable of breathing both in water and in air. Their morphology suggests that they fed on a variety of kinds of foods. Some forms like Mixopterus were very scorpion-like and may have even been ancestral to scorpions. About two dozen families of eurypterids are known.
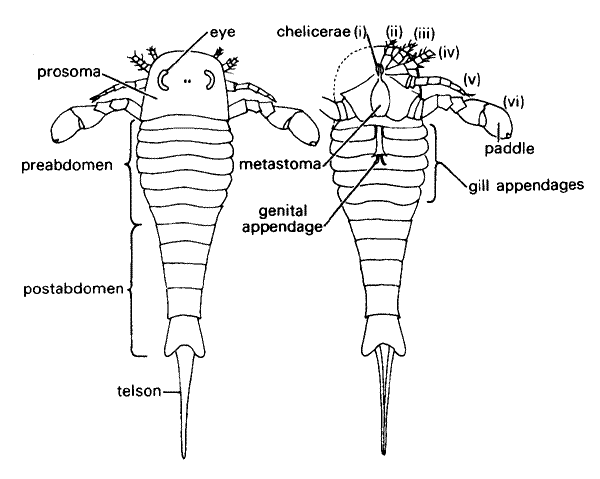
Baltoeurypterus tetragonophthalmus
left, dorsal (upper) view; right ventral (bottom) view
The morphology of the Eurypterid exoskeleton is very well known, thanks to the work of Holm in 1898, who made a detailed study of the Silurian species Baltoeurypterus tetragonophthalmus, formerly known as Eurypterus fischeri. He did this by isolating fragments of the exoskeleton from rock, as he did with graptolites, so that they could be studied as transparencies.
The eurypterid body is similar to that of other primitive arthropods, such as trilobites and horse-shoe crabs (Xiphosura). In fact the Eurypterids are rather closely related to the latter. As with the Xiphosura, the long segmented body can be divided into two parts.
The front or "head" part is called the prosoma, where the legs are attached and the eyes and mouth are located. As with horseshoe crabs, the prosoma is both the head and thorax of the animal. The upper or dorsal side of the prosoma is equipped with two prominent kidney-shaped compound eyes, and in the middle a pair of small ocelli or simple eye-like sense-organs. The mouth is located underneath in the center, and around it the six pairs of legs are attached. Depending on their structure, these were used for walking, swimming, or grasping food. The legs are connected to the body by toothplates or gnathobases. These border the slit-like mouth and are lined by teeth.
The first pair of legs are the chelicerae, which are the same organs (but differently developed - i.e. homologous) as the fangs of spiders. All chelicerate arthropods have this special first set of modified legs. The next four pairs of appendages are the walking legs, These are cylindrical and spiny and increase in size with each pair. Finally there are the very large smooth flattened paddle-like sixth pair of limbs, which were used for swimming. These were capable of being protracted and retracted and were probably used in a breast-stroke-like manner. Note that there are no antennae - Chelicerates do not have antennae. A small U-shaped plate (the endostoma) borders the mouth; and is normally covered by a much larger plate, the metastoma, which is actually part of the abdomen
The long rear part is the opisthosoma, which can be further divided into a broad flattened pre-abdomen consisting of seven segments, and a narrower and more cylindrical post-abdomen of only five segments. Alternatively it can be divided into the first six (the mesosoma - "middle body") and the last six (metasoma). All Eurypterid species have a total of twelve flexible opisthomal segments, or tergites as they are called. At the end of the last segment is a stout blade-like spine, the telson.
The prosoma is actually made up of the first six exoskeleton segments, which have been fused together. The seventh segment has become the metastoma; and the eighth is the genital aperture. Xiphosurans have this same genital aperture but do not have a metasoma. This plate-like eighth segment is also termed the operculum. In its center is the genital appendage, an elongated and sculptured rod of which there are two kinds always found in a single eurypterid population. Traces of internal ducts have been found in both kinds.
Each of the abdominal segments is composed of a dorsal (upper or back) tergite and a ventral (lower or belly) sternite. The operculum and the following four appendages of the pre-abdomen covered chambers in which the five pairs of gills were located. These were covered by protective plates. The first pair of gills/plates also contain the reproductive organs. In some forms the gills were able to remain moist and so allow the animal to move about on land for short periods
Eurypterid cuticle was chitinous and of varying thickness, but generally very thin, and preserved specimens (which were probably mostly molted skins) are usually crushed. It may bear different kinds of sculpture, such as terrace lines and scale-like ornament. Ultramicrographic work has revealed inner laminations resembling those of Limulus the modern horseshoe crab.
Not much is known about the internal organs of eurypterids. There are a few fossils in which part of the intestine is preserved, and this ends in an opening between the final tergite and the telson. It is not unlikely that the internal structure of Eurypterids would have been midway between that of their two closest living relatives - the horseshoe crab and the scorpions. Recently more has been known thanks to exceptionally preserved material from the Late Ordovician of South Africa, displaying internal tissues such as the musculature associated with the appendages and telson. Also, material from the earliest Carboniferous (early Tournasian) of France reveals a possible endosternite, gut trace and lamellate gills.
It was originally suggested, e.g. by Størmer, an important authority on the Merostomata, that Eurypterids flipped over and swum on their backs, using their gill plates like huge fairy shrimps.
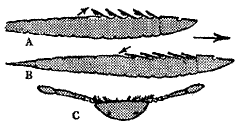
Upside-down swimming, possibly used by tiny young Eurypterids, but not by adults
(from Moore, Lalicker and Fischer, Invertebrate Fossils)
This explanation (see figure above) is unlikely in view of the upward-facing eyes, and recent studies have shown that swimming involved using the large last set of limbs as a paddle or oar.
However, it is still possible that young individuals may have practiced upside-down swimming, as young xiphosurans do today.
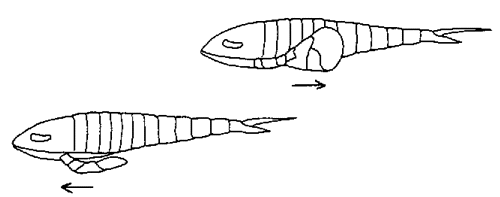
Baltoeurypterus swimming
from Clarkson's Invertebrate Paleontology and Evolution, 3rd ed.
Although many Eurypterids had legs too tiny to do more than allow them to crawl over the sea bottom, a number of forms had large stout legs, and were clearly capable of terrestrial locomotion (like land crabs today). While functional studies that eurypterids used out-of-phase walking techniques, their trackways indicate that they used in-phase, hexapodous (six-legged) and octopodous (eight-legged) gaits.
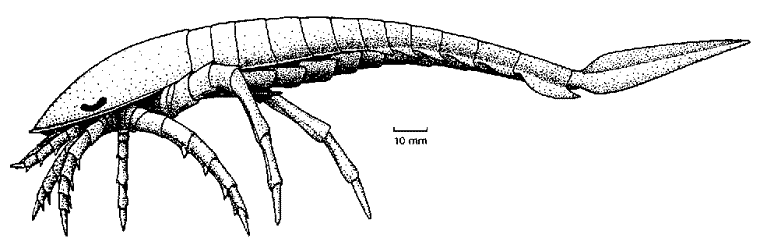
Parastylonurus walking
from Clarkson's Invertebrate Paleontology and Evolution, 3rd ed.
Understanding sexual dimorphism among extinct organisms is one of the many problems that bedevil paleontologists in their attempt to reconstruct organisms known only from bones, shells, or carapaces. In some cases the two sexes of an organism have been interpreted as two distinct species living side by side; in other instances there is not enough difference to even distinguish different sexes. And even when they can be differentiated, there is the problem of figuring out which is the male and which the female.
The eurypterids present a particularly difficult example of this. As mentioned, there are always two kinds of genital appendages found in any Eurypterid population. The original hypothesis has the type A, which is equipped with "clasping organs" on the prosomal appendages, as the male copulatory organ, while the other kind (type B) can be interpreted as the female ovipositor for depositing eggs, and fitting neatly over the male clasper. However, Simon Braddy suggests that eurypterid reproduction occurred via spermatophore (sperm package) transfer on the substrate (in at least Baltoeurypterus); the type A appendage is in this case the female who is able to retrieve a spermatophore for storage in her spermathecae ('horn organs'). The 'scimitar lobes' (male) are considered to have been clasping structures.
In any case the variations of form among different taxa are very useful in classification. To avoid the possibility of error, I have simply referred these to "type A" and "type B", leaving open which type belongs to which gender.
Eurypterids inhabited marine, brackish, and freshwater environments. Most were active benthos, although some were nektonic, being able to actively swim. Some may have been able to spend short intervals of time on land. Most eurypterids were probably predatory on other eurypterids or fishes. Eurypterid associations can often be distinguished and related to environmental conditions, such as marine, lagoonal or estuarine, or brackish to freshwater faunas.
Dr Simon Braddy rejects Kjellesvig-Waering's 'ecological phases' model in favour of a model whereby individual taxa have a palaeoecological range influenced by an 'ontogenetic gradient'. A 'mass-moult-mate' hypothesis, comparable to the behavior adopted by some extant semi-terrestrial crustaceans, thus explains the occurrence of abundant eurypterid assemblages (e.g. the Bertie Waterlime), previously regarded as mass death assemblages.
The first certain eurypterids appeared during the Middle Ordovician, although a possible earlier ancestral form (Kodymirus, although this may be an aglaspid) dates back to the Early or Middle Cambrian. They seem to have diversified quite quickly, and by the end of the Ordovician almost all the main lineages had appeared. Although the morphology of some genera is well known others are represented only by very poorly preserved material, and there are acute taxonomic problems in classifying such isolated remains. Differences involve adaptive differences in size, prosomal shape, the location of the eyes, and the morphology of the body, the appendages and the telson. The best know classification is the Linnean arrangement given in the Treatise on Invertebrate Paleontology, which is now sadly out of date. Dr Simon J. Braddy uses a cladistic analysis of the more completely known eurypterid genera (48 taxa, 19 characters) to give the following model of eurypterid phylogeny:
The classification given on the following pages is based on a synthesis of Treatise on Invertebrate Paleontology, The Fossil Record I (I don't have the revised edition unfortunately), Clarkson's Invertebrate Paleontology and Evolution, and the above cladogram of Dr Simon J. Braddy, updated with a few other recent web sources which appear to have some reliability. Combining as it does a number of sources, and as I am unable to acquire more detailed up to date information at the moment, this classification should be taken as being very provisional only. Click on each image and group for more details.
|
Suborder Eurypterina |
Suborder Stylonurina |
||
 |
 |
 |
|
| Eurypteracea | Mixopteracea | Pterygotacea |
Stylonuracea (see Stylonuridae) |
Eurypterida Burmeister 1943
Synonyms Gigantostraca Haeckel 1866
Ordovician to Permian, Cosmopolitan
Phylogeny: Chelicerata : Xiphosura + (Arachnida + * : Stylonuracea + (Eurypteracea + (Mixopteracea + -Pterygotina)
Links: Eurypterida - UCMP; The possible lifestyle of a Late Ordovician eurypterid - by Jeffrey Minicucci - very interesting essay on the mode of life of Onychopterella augusti; Faktablad om sjøskorpioner - material on Eurypterids - in Norwegian - includes a photo of a lovely Mixopterus specimen; Eurypterida - Dendrogram and genus list, by Mikko K. Haaramo; Dr Simon J. Braddy - research on Eurypterids, and on Paleozoic arthropod trackways as documenting the diversity and palaeoenvironmental distribution of the early terrestrial arthropods.; bulletin board Re: Eurypterid Chat; Where? - some notes on the late Silurian Williamsville formation of Buffalo, New York, and the Eurypterid fauna
page uploaded 12 June 2002
(originally uploaded on Kheper Site 22 February 1999)
checked ATW040206
text by M. Alan Kazlev Creative Commons Attribution 1999-2002
last modified ATW060226
