Arthrodira
Taxa on This Page
- Arthrodira X
The Arthrodires -- Introduction
The name Arthrodira means "jointed neck" These advanced preditory placoderms had a movable joint between the head cephalic) and the thoracic (body) armor (you can see the way the armour is divided into two halves in the above photo). The mouth is interesting because as the lower jaw moved down the head shield moved up, thus giving a wide gape. Like all placoderms, they lacked teeth, instead using the sharpened edges of a bony plate as a biting surface.
There is a misconception that these creatures were sluggish bottom-dwellers. But as discussed below concerning this successful group, the Arthrodires were not only one of the most diverse and numerically sucessful vertebrate clans of the Devonian, but they occupied or even dominated virtually every ecological guild which would admit vertebrates, from top predator to scum-sucking bottom-dweller, for about fifty million years.
Arthrodires changed little during their long (50 million years is only slightly less than the time mammals have ruled the Earth), although during the end of their reign on lineage did increase enormously in size, perhaps exploiting the food source provided by diversifying sharks and bony fish. The largest genus of this group, Dunkleosteus (see illustration above), was a true superpreditor of the latest Devonian (Famennian epoch), reaching 3 to as much as 9 meters in length. The mighty head-shield alone (see above photo) was upto a meter in height in large forms. Note the sharpened edges of the jaw, which functioned as teeth. Like other sharp-sighted animals (birds, ichthyosaurs, some archosauriforms and dinosaurs) the eye sockets are protected by a bony ring.
The arthrodires were not out-competed by more advanced fish, as is usually but incorrectly believed. Rather they were eliminated by a terminal devonian extinction event, allowing the sharks to diversify into the vacated ecological niches during the Carboniferous period. MAK
More on Arthrodires
Placoderms make an interesting and useful study because there is little danger of being misled by living species. The closest living relatives of the Arthrodires are lampreys, in one direction, and holocephalians in the other - which suggests just how far in the taxonomic wilderness this group is. There is never a lack of unanswered questions in paleontology, but the placoderms do seem to have more than their fair share. Only a very few of these questions are addressed here, and certainly none are answered.
Some scholars seem to have consigned the placoderms to a slow, dim life as bottom-feeders, scraping by on detrital scraps fallen from the tables of the evolving sharks and actinopterygian ray-fin fish who whizzed above them like F-14s passing a garbage truck. This general concept of the placoderms cannot survive contact with the Arthrodira. The arthrodires were not only one of the most diverse and numerically successful vertebrate clans of the Devonian, but they occupied or even dominated virtually every ecological guild which would admit vertebrates, from top predator to scum-sucking bottom-dweller (both positions are, of course, now occupied by lawyers) for about fifty million years. There are altogether too many illustrations of the 3 meter Dunkleosteus chasing the 50 cm shark, but they do make a valid point.
Why was the dentition so conservative? After 50 My, the last arthrodire had two superognathal plates and one inferognathal, not terribly different from the first. Even the dentally unimaginative dinosaur clades often did better than this. No placoderm ever developed teeth from scales in the way of other vertebrate groups, and it is unclear how, if at all, placoderms replaced damaged dentition. The form of the gnathal plates seems to have been subject to considerable variation. Some had sharply se cutting surfaces, some had broad crushing surfaces, others had little or nothing recognizable. However, the basic gnathal formula was extraordinarily stable.
The rigid head and body armor is also unique among gnathostomes, but is perhaps more easily explained. The reader is cautioned that the following argument may be reasonable, but is entirely speculative at present. The vertebral support of the placoderms, in fact almost all Devonian gnathostomes of all groups, is fairly weak. Strong vertebral support is obviously less critical for aquatic organisms. However, strong longitudinal support allows stronger muscle movements in swimming than a notochord might permit. Furthermore, aguilliform eel-like) movement excepted, efficiency of the notochord as a device for storing and directing the energy exerted by swimming muscles goes down strongly as a function of body length; nor is it possible to simply increase the diameter of the notochord as the diameter of the animal expands. [There, may, in addition, be a variant of the classic square-cube problem] There is only so much one can do with a rod of hydrated gristle.
The placoderms solved these problems in a very straightforward manner -- by evolving an exoskeleton, or at least by retaining an exoskeleton that their eugnathostome relatives had been so foolish as to squander. To be more exact, they evolved half an exoskeleton. The other half of the body remained free to make extremely strong swimming motions, anchored on the rigid craniothoracic exoskeleton without the body as a whole being forced to subscribe more than one half of a sinusoidal wave (this being, roughly, what makes a tuna fast and an eel slow but "slippery" and agile). The posterior of placoderms, where known, is thinner than expected for the equivalent osteichthyan or shark and generally scale-less, although there are exceptions. Quite likely this was because its principal function was to exert motive power -- more along the lines of a gigantic flagellum than an extension of the body cavity as such. Undoubtedly, it made better sense to attach the internal organs to the reinforced exoskeleton, near the center of gravity and of rotation, rather than dispersing them along the trunk in the manner of modern fish or tetrapods.
The posterior trunk of placoderms is also surprisingly finless. Again, our knowledge of this anatomical region is limited. However, there is no evidence of a substantial dorsal fin or of anal fins in any arthrodire, and little enough evidence of caudal fins. The primary control surface was necessarily the pectoral fin. The oddly jointed pectoral fin armor is a memorable feature of the placoderms, especially Antiarchs. It is sometimes assumed that this armor was the fin. However, it may perhaps be better understood as the exoskeleton of a very large and powerful control surface. Recall that the head of most arthrodires was capable only of limited movement in the vertical plane, and none at all horizontally due to the accessory attachments on the exoskeleton. The movement of plates relative to each other was also minimal (recall the lack of overlap). The only possible control surface is the pectoral fin.
Thus, paradoxically, the arthrodires and, more generally, the placoderms may have succeeded because they were specialized for speed and for an active, pelagic lifestyle. They may, in fact, have overspecialized. Later arthrodires show some signs of convergence with osteichthyans, before the entire group was eliminated for an unknown reason towards the end of the Devonian. However that may have been, it cannot be assumed that the arthrodires were merely the leading clan in a caste of benthic snails. ATW
Descriptions
Arthrodira: Coccosteus, Dicksonosteus, Dunkleosteus.
Early Devonian to Late Devonian
 |
| Dunkleosteus armour from The Devonian Period -- Pamela J. W. Gore |
Highly diverse and successful group in various roles, including dominant marine predator. $ Two sets of upper tooth plates (superognathals); gnathal plates may be tuberculated; large endocranial postorbital process; $ skull roof from 3 unpaired medial plates + 7 paired plates; cranial plates do not overlap; 3 cheek plates; hyomandibula fused w. cheek, not part of jaw support; inferognathal typically lies on Meckelian bone, with adductor muscles apparently running directly between Meckelian and palatoquadrate; anterior braincase with nasal capsules separated by perichondrally ossified "wall", the cranio-ethmoid fissure, which in some species completely separates nasal capsules from brain; olfactory capsules large and open ventrally (just ant to superognathals); cartilage eye-stalk links ball to braincase through (sometimes ossified) cup-shaped sclera; orbit hollowed with myodomes, especially posterior; palatoquadrate articulates with ethmoid and with dermal armor; braincase also articulates with gill arches; sensory line system in pits or grooves on dermal armor; $ endolymphatic opening in single paranuchal plate; main joint between braincase and synarcual; extra joint between head and body armor in most (but not Actinolepidoids) with condyles on anterior dorsolateral plate of thoracic armor & fossa on paranuchal plate of head armor (note that this stabilizes the head but prevents any significant lateral motion); primitively, elongated thoracic armor, with dermal plate closure posterior to pectoral; thoracic plates have small overlap; axial skeleton otherwise unknown; pectoral fin endoskeleton unknown, but scapulocoracoid shows narrow base for fin (stenobasal), with muscle attachments and vascular canals attaching external to base; some degree of pectoral armor common; dermal armor thick, cellular bone with external tubercles of semidentine absent in some species which may have been skin-covered). (MAK 991218)
Image: from Science -- Burrow 300 (5626)- 1661b. Cross section of an 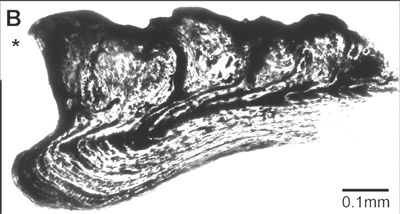 arthrodire gnathal plate. See comments below. To us, these look about halfway between the conjoined denticles sometimes found internally in thelodont gills and gnathostome scales.
arthrodire gnathal plate. See comments below. To us, these look about halfway between the conjoined denticles sometimes found internally in thelodont gills and gnathostome scales.
Links: devonian image page (Henderson's Dunkleosteus); Fakstasider Norwegian); Carolowilhelmina; Arthrodira Mikko's Phylogeny); Geotimes - April 2003 - Devonian Dentistry non-technical abstract of very important new finding -- Arthrodires had teeth!); Sciscape 新聞 [Mar 03, 2003]: 不只一次--牙齒的起源與演化 Chinese version of same article); ScienceNow another summary, with some cogent academic comment); Science -- Burrow 300 (5626)- 1661b an unconvinced commentator); Science -- Smith and Johanson 300 (5626)- 1661c response by original authors); UN ARTHRODIRE BRACHYTHORACI (PLACODERME) French article describing a new, fragmentary arthrodire); Robert K. Carr some links & diagrams); Placodermi, (Klass Gnathostoma) Swedish: general information). ATW030703.
checked ATW0151003; new page MAK111027

