Archosauria: Rauisuchiformes: Prestosuchidae
Saurosuchus
Taxa on This Page
- Prestosuchidae X
- Saurosuchus X
Prestosuchidae
Included here is the beginning of an assemblage of rather poorly known but phylogenetically important crurotarsan archosaurs. The Prestosuchidae probably fall somewhere in the middle range of basal Archosauria. In most cladograms they more derived than phytosaurs and ornithosuchids, but usually less derived than the rauisuchids and aetosaurs of the Late Triassic. The prestosuchids were active terrestrial predators, and were highly successful during the Triassic. These were all large animals, even the smaller representatives being around 2.5 meters in length. Saurosuchus reached 6 metres and more.
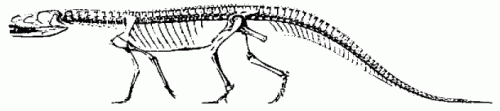
|
|
Ticinosuchus ferox, from the Grensbitumenhorizonts Anisian/Ladinian boundary, middle of the Middle Triassic) of Monte San Giorgio, Tessin Alps, Switzerland. Reconstructed skeleton, length about 2.5 meters. AH Müller, Lehrbuch der Paläozoologie, p. 296, after Huene (1956).
|
The Prestosuchidae have sometimes been included in the Rauisuchidae, which are variously considered the sister group of the Crocodylomorpha (Gauthier, 1986), the Ornithosuchidae (Benton & Walker, 2002), or the sister group of the aetosaurs, the two constituting a monophyletic Pseudosuchia (Benton & Clark, 1988). Alternatively, separate from the more advanced Rauisuchia, they are variously referred to as an independent lineage that falls outside a crocodylomorph - poposaurid - rauisuchid - aetosaur clade Parrish, 1993), or the sister group of the aetosaurs in a monophyletic Pseudosuchia Juul, 1994). However, most investigators find that the synapomorphies (unique features) that characterize the prestosuchids are not enough to place them securely in a separate clade. So, it is entirely likely that they represent a paraphyletic assemblage or gradation (like, say, "Pelycosauria" or "Therapsida") that are intermediate between primitive forms (in this case basal Crurotarsi) and more advanced types (such as the Aetosauridae and Rauisuchidae), with the Prestosuchidae proper being a much smaller assemblage (Gower 2002). In our hands, they fall in a fairly neat morphocline between the ornithosuchids and rauisuchids.
The success of these animals during the Triassic may have been due in part to their reptilian (sauropsid) metabolism which gave them an edge over our synapsid relatives in the arid Pangean climate, and in part to their efficient stance. Unlike earlier archosaurs, which had a typically reptilian sprawling or only semi-erect limb-posture, the "prestosuchid" archosaurs, and the rauisuchids and poposaurids (postosuchids) had a fully erect gait and posture. They had mobility like dinosaurs and mammals, rather than like reptiles. Benton (1984). Increasingly efficient limb-posture and mobility may also indicate that these creatures had a different, perhaps more energetic, metabolism than their predecessors. Their main centre of evolution seems to have been West Gondwana, although a relatively small form (Ticinosuchus) flourished in Europe during the mid-Triassic.
Anatomically, the prestosuchids are a more massive, solid version of the ornithosuchids. The cranial peculiarities of Ornithosuchus discussed previously are still present, but sometimes masked by the greater thickness of bone. The protruding premaxilla, still partly separated from the maxilla has become even more blocky and rectangular, particularly in Saurosuchus. The basic structure of the antorbital fenestra is the same, although the twisted antorbital bar has been replaced by a straight, girder-like extension of lacrimal which locks on to a jugal process oriented at a right angle to the lacrimal process. The prefrontal "awning" is smaller, but still present, and is elongated by a similar lateral process of the frontal which extends laterally over the orbit. The region around the upper temporal fenestra looks even more like Sinraptor than does the corresponding region in Ornithosuchus. In fact, when due allowance is made for a heavier covering of bone, there is little to distinguish the dermal skulls of Ornithosuchus and Saurosuchus. The palate is more distinctive, but the prestosuchid skull is much wider, and the anterior palate of Ornithosuchus is not well known, so that one cannot make to much of the comparison. The postcranial skeleton is likewise quite similar, again with due allowance for the difference in mass. The pelvis, in particular, is built along very much the same lines.
MAK 991019 and 030808. ATW040130.
Descriptions
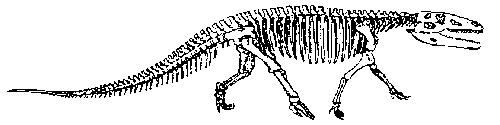
|
| Prestosuchus sp., from the Carnian of Brazil (Santa Maria Fm); reconstructed skeleton, length about 5 meters. A. H. Müller, Lehrbuch der Paläozoologie, p.296. after Huene 1956 |
Prestosuchidae: Prestosuchus chiniquensis Huene, 1942, Prestosuchus loricatus Huene, 1942, Tikisaurus romeri Chatterjee & Majumdar, 1987.
Range: Late Middle and Early Late Triassic of Gondwana Ladinian & Carnian of South America; Late Carnian of India).
Phylogeny: Rauisuchia ::: Doswellia + (Rauisuchidae + Poposauridae) + * : Saurosuchus + (Ticinosuchus + Batrachotomus).
The Prestosuchidae have previously been included under the Rauisuchidae, or as a large Middle and Late Triassic grouping, but are here defined as a much smaller clade outside the Suchia
 Characters: medium to large size [S74]; skull large, narrow & deep [S74]; some genera with small crescentic accessory antorbital fenestra (probably homologous with max-premax diastema of ornithosuchids, thus a rauisuchian synapomorphy) [S74] [π$03]; antorbital fenestra large & elongate [S74]; upper temporal fenestra small [S74]; descending process of prefrontal present [π$03]; orbit keyhole-shaped [S74]; postorbital-jugal bar with distinct "step" [π$$03]; exoccipital with lateral crest lying anterior to hypoglossal foramen [$G02] [π$$03]; ventral ramus of opisthotic extends laterally further than exoccipital [π03]; foramina for cerebral branch of carotid on lateral face of basisphenoid [G02] [1]; maxillae meet at midline anterior to choana to form secondary palate [π$03]; teeth recurved, flattened & serrated [S74]; neural arches high, not inclined, rectangular, with deep interspinous notch [S74]; accessory neural spine on midcaudal vertebrae [π$03] [3]; manus V with 4 phalanges [π$$03]; pelvis triradiate [S74]; ilium with strong posterior process & brevis shelf [S74]; ischium elongate & rod-like, fused at midline along most of its length [S74]; pubic plate reduced and pubis with little participation in acetabulum [S74]; pubis shorter than ischium (reversal) [π$03]; acetabulum closed [S74]; acetabulum ventrally deflected [π$03] [4]; femur long & slightly sigmoid [S74]; femur 4th trochanter poorly defined or absent [S74]; shallow socket on the calcaneum articulating with astragalar ball ("crocodile-normal" tarsus) medial to the fibular facet [S74] [$P93]; posteriorly directed calcaneal tuber [J Conrad, iNet]; calcaneal tuber, distal end with dorsoventrally aligned median depression [π$$03]; metatarsal V hooked [S74]; pes V with 1 phalanx [π$$03]; erect limb posture; quadrupedal [S74]; paramedian plates with anterior articular processes [$?P93] and posterior dorsal keel [S92]; carnivorous.
Characters: medium to large size [S74]; skull large, narrow & deep [S74]; some genera with small crescentic accessory antorbital fenestra (probably homologous with max-premax diastema of ornithosuchids, thus a rauisuchian synapomorphy) [S74] [π$03]; antorbital fenestra large & elongate [S74]; upper temporal fenestra small [S74]; descending process of prefrontal present [π$03]; orbit keyhole-shaped [S74]; postorbital-jugal bar with distinct "step" [π$$03]; exoccipital with lateral crest lying anterior to hypoglossal foramen [$G02] [π$$03]; ventral ramus of opisthotic extends laterally further than exoccipital [π03]; foramina for cerebral branch of carotid on lateral face of basisphenoid [G02] [1]; maxillae meet at midline anterior to choana to form secondary palate [π$03]; teeth recurved, flattened & serrated [S74]; neural arches high, not inclined, rectangular, with deep interspinous notch [S74]; accessory neural spine on midcaudal vertebrae [π$03] [3]; manus V with 4 phalanges [π$$03]; pelvis triradiate [S74]; ilium with strong posterior process & brevis shelf [S74]; ischium elongate & rod-like, fused at midline along most of its length [S74]; pubic plate reduced and pubis with little participation in acetabulum [S74]; pubis shorter than ischium (reversal) [π$03]; acetabulum closed [S74]; acetabulum ventrally deflected [π$03] [4]; femur long & slightly sigmoid [S74]; femur 4th trochanter poorly defined or absent [S74]; shallow socket on the calcaneum articulating with astragalar ball ("crocodile-normal" tarsus) medial to the fibular facet [S74] [$P93]; posteriorly directed calcaneal tuber [J Conrad, iNet]; calcaneal tuber, distal end with dorsoventrally aligned median depression [π$$03]; metatarsal V hooked [S74]; pes V with 1 phalanx [π$$03]; erect limb posture; quadrupedal [S74]; paramedian plates with anterior articular processes [$?P93] and posterior dorsal keel [S92]; carnivorous.
Notes: [1] synapomorphy of clade uniting Prestosuchidae with Suchia [G02]. [2] "[π03]" refers to our own analysis of published data. The prefix "$$" indicates a synapomorphy of the larger clade including Prestosuchidae + Rauisuchidae. [3] Might be related to supposed angled mid-caudal vertebral centrum of Doswellia. [4] homologous to, but less extreme, than condition in Postosuchus.
Links:
Rauisuchia Translation and Pronunciation Guide Introduction.
Reference: Gower 2002 [G02], Parrish (1993) [P93], Sill (1974) [S74], Sues (1992) [S92]. ATW031213.
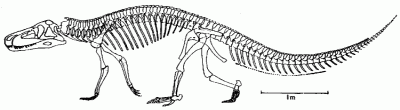
|
|
S. galilei, from the Carnian of Argentina (Ischigualasto Formation); reconstructed skeleton (scale bar 1 meter). From Benton (1984).
|
Saurosuchus: Saurosuchus galilei Reig 1959
Range: Late Triassic (Carnian) of South America Argentina).
Comments: Saurosuchus, known from a skull and several postcranial remains, from both the lower and the upper sections of the Ischigualasto Formation, San Juan Province, Argentina, was an enormous predator. With a length of 6 or 7 metres and a skull alone of up to a meter in length, it was as large as a decent-sized Jurassic or Cretaceous theropod (say a medium-sized Allosaurus or 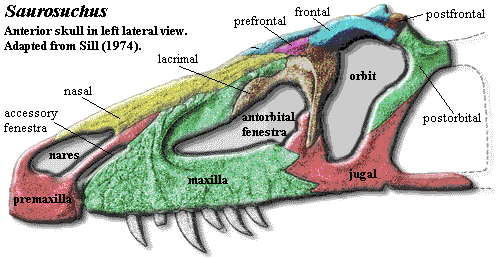 tyrannosaur). Although in some features primitive [P93] [G02] it was more advanced than Prestosuchus and Rauisuchus, with a more elongate ischium and femur, and strong pneumatisation (air-spaces) of the vertebrae [B71], all of which would seem to indicate a more active and metabolically advanced animal. Without doubt the "top predator" of the day, this huge carnivore would have fed upon the large kannemeyeriid dicynodonts whose remains occur in the same formation.
tyrannosaur). Although in some features primitive [P93] [G02] it was more advanced than Prestosuchus and Rauisuchus, with a more elongate ischium and femur, and strong pneumatisation (air-spaces) of the vertebrae [B71], all of which would seem to indicate a more active and metabolically advanced animal. Without doubt the "top predator" of the day, this huge carnivore would have fed upon the large kannemeyeriid dicynodonts whose remains occur in the same formation.
Characters: large (to 6m) [S74]; skull deep, transversely narrow, elongate & strongly constructed [S74]; skull roof finely sculptured [S74] [A00]; body of premaxilla is massive rectangular block with 2 narrow processes on either side of naris [S74] [A00]; premaxilla with thickened oral rim bearing 4 tooth positions [S74] [A00]; premaxillae with long, thick symphysis [S74]; premaxillae not strongly sutured to maxilla [S74]; very large nares bordered only by nasals & premaxillae [S74]; accessory subnarial fenestra between maxilla and premaxilla [S74] [$P93]; maxilla U-shaped, large & plate-like, forming entire ventral margin of antorbital fenestra & half of dorsal margin (overlapping part of lacrimal) [S74] [A00]; maxilla forms prominent crista parallel to ventral border of fenestra [A00]; maxilla with large foramen on medial surface, just anterior to jugal suture [S74]; maxilla sculptured [A00]; antorbital fenestra large (slightly smaller than orbit) & elongate, with well-defined smooth border [S74]; nasal anteriorly thin, very elongate, overlapping medial
 process of premaxilla [S74] [A00]; nasal thickens & becomes somewhat broader posteriorly [S74] (contra [A00]); nasal with ornament of irregular anteroposterior grooves [S74]; prefrontal small, oriented dorsally & not contacting orbit [S74] (contra [A00], participates in orbit); prefrontal with distinct posterior process underlying frontal dorsal to orbit, joining with parietal to form ventral depression having possible kinetic contact with laterosphenoid [A00] [π$03] [3]; frontal dominated by massive supraorbital arch [S74] [A$00]; frontal with pit & groove ornament [S74]; frontal with median longitudinal ridged (nasal tract?) [S74]; frontal with pocket posteriorly for laterosphenoid anterior process [S74]; frontal with lateral process forming posterolateral margin (i.e. the supraorbital arch projects laterally) [A$00]; upper temporal fenestra small & on dorsal face of skull [S74]; postfrontal reduced, not participating in orbit & hidden in dorsal view by frontal [A$00]; parietal forms dorsal border of upper fenestra [A00]; parietal overlaps supraoccipital and contacts laterosphenoid [A00]; parietal crest which runs along posteromedial border of upper fenestra, participates in posttemporal foramen, & broadly overlaps squamosal [A00]; orbit keyhole-shaped [S74]; lacrimal lies mostly in antorbital fenestra as smooth-surfaced thin plate [S74]; lacrimal forms cylindrical process and prong posteriorly, overlying antorbital process of jugal in a probably mobile articulation [S74] [A$00]; possible lacrimal foramen near border with frontal [S74] (contra [A00] no foramen); lacrimal strongly sutured to prefrontal [A00]; lacrimal without transverse component [S74]; strong orbital arch formed by frontal [S74]; jugal having zig-jag suture with maxilla [S74] (contra [A00] jugal with moveable, overlapping contact with maxilla); jugal with pit & groove sculpturing [S74]; jugal with strong diagonal abutment with postorbital [S74]; jugal with thin, tapering bar below lower fenestra [S74] [A00]; jugal with
process of premaxilla [S74] [A00]; nasal thickens & becomes somewhat broader posteriorly [S74] (contra [A00]); nasal with ornament of irregular anteroposterior grooves [S74]; prefrontal small, oriented dorsally & not contacting orbit [S74] (contra [A00], participates in orbit); prefrontal with distinct posterior process underlying frontal dorsal to orbit, joining with parietal to form ventral depression having possible kinetic contact with laterosphenoid [A00] [π$03] [3]; frontal dominated by massive supraorbital arch [S74] [A$00]; frontal with pit & groove ornament [S74]; frontal with median longitudinal ridged (nasal tract?) [S74]; frontal with pocket posteriorly for laterosphenoid anterior process [S74]; frontal with lateral process forming posterolateral margin (i.e. the supraorbital arch projects laterally) [A$00]; upper temporal fenestra small & on dorsal face of skull [S74]; postfrontal reduced, not participating in orbit & hidden in dorsal view by frontal [A$00]; parietal forms dorsal border of upper fenestra [A00]; parietal overlaps supraoccipital and contacts laterosphenoid [A00]; parietal crest which runs along posteromedial border of upper fenestra, participates in posttemporal foramen, & broadly overlaps squamosal [A00]; orbit keyhole-shaped [S74]; lacrimal lies mostly in antorbital fenestra as smooth-surfaced thin plate [S74]; lacrimal forms cylindrical process and prong posteriorly, overlying antorbital process of jugal in a probably mobile articulation [S74] [A$00]; possible lacrimal foramen near border with frontal [S74] (contra [A00] no foramen); lacrimal strongly sutured to prefrontal [A00]; lacrimal without transverse component [S74]; strong orbital arch formed by frontal [S74]; jugal having zig-jag suture with maxilla [S74] (contra [A00] jugal with moveable, overlapping contact with maxilla); jugal with pit & groove sculpturing [S74]; jugal with strong diagonal abutment with postorbital [S74]; jugal with thin, tapering bar below lower fenestra [S74] [A00]; jugal with 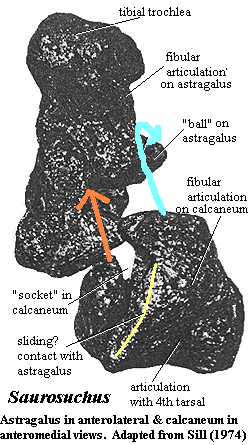 medial bulge just ventral to orbit marking internal, kinetic ectopterygoid contact [S74] [A00]; postorbital triradiate, fairly large, with prominent tuberosity just posterior to orbital arch [S74] [A00]; postorbital forming lateral and anterior border of upper temporal fenestra [S74] [A00]; postorbital with short posterior process fitting under anterior process of squamosal [A00]; lower temporal fenestra approximately same size as orbit [S74]; squamosal with ventral process overlapping quadratojugal [A00]; squamosal with short, blunt posterior process [A00]; quadrate vertical with thick vertical column bearing anterior flange which overlaps quadratojugal [A00]; quadrate column curved posteriorly for to form border for tympanum [A00]; quadrate with convex articular condyle [A00]; supraoccipital contributes to foramen magnum [π$03]; supraoccipital triangular, fused to exoccipitals & opisthotics [A00]; exoccipitals fused to opisthotics and paroccipital process [A00]; paroccipital process not fully ossified distally [A00]; metotic foramen communicates nerves IX-XI, with separate hypoglossal foramen posterior to metotic foramen [A00]; paroccipital process forms base of posttemporal foramen [A00]; basioccipital forms most of occipital condyle [A00]; condyle bearing notochordal pit [A00]; basioccipital broadly overlaps basisphenoid [A00]; basioccipital with well-developed basal tubers [A00]; basioccipital with blind basioccipital recess [A00]; semilunar depression absent from braincase [G02]; basisphenoid dorsoventrally short [G02]; presphenoid may [A00] or may not [G02] be present above trigeminal notch; olfactory bulb at end of long peduncle [A00]; maxilla with palatal process massive, meeting on midline and vaulted, meeting vomer (also contact with palatine & ectopterygoid) [S74] [A00]; long triangular interpterygoid vacuity present [S74]; vomers not contacting premaxilla (but see figure) [A00]; palatines thin, with overlapping contacts [A00]; palatines, and palate generally, arches upward sharply [A00]; pterygoid large, contacting, vomer, ectopterygoid, palatine, quadrate & basisphenoid, bearing the usual palatal, transverse and quadrate rami [A00]; anterior ramus strongly convex ventrally & near vertical at midline [A00]; ectopterygoid with expanded lateral foot contacting jugal [A00]; dentary robust, not dorsoventrally expanded, & external surface is almost flat [A00]; dentary medial surface almost covered by splenial & coronoid [A00]; dentary symphysis with two flanges grasping opposite ramus [A00]; palatal dentition absent [S74] [P93]; teeth homodont, large, robust, laterally compressed, recurved, sharply pointed & serrated [S74]; tooth roots twice as long as crowns [S74]; teeth with thin, striated enamel [S74]; 4 teeth on premaxilla, 10 on maxilla [S74] (contra [A00] 12 on maxilla); interdental plates present [S74] [A00]; vertebrae amphicoelous [S74]; vertebrae with neural spines anteroposteriorly elongate & flat [S74]; vertebrae with prominent interspinous notch on anterior face [S74]; vertebrae with grossly constricted centra, leaving thin, vertical plate between flared rims [S74]; 23-25 presacral vertebrae [S74]; dorsal vertebrae compressed laterally & reduced to thin vertical plate between flared ends [S74]; rib facets well separated & on different levels throughout column, but never on centra [S74]; transverse processes short [S74]; prezygapophyses short, not overhanging centrum [S74]; neural spines flat, tall, thin, not inclined [S74]; ribs massive, with prominent ridge on proximal anterior surface [S74]; 2 sacral vertebrae, not fused & generally similar to dorsals [S74]; 2nd sacral & 1st caudal with anterior centrum flare enclosing and immobilizing end of preceding centrum [S74]; sacral transverse processes massive [S74]; caudal vertebrae shorter & with large rounded rims, less strongly constricted than dorsals (&
medial bulge just ventral to orbit marking internal, kinetic ectopterygoid contact [S74] [A00]; postorbital triradiate, fairly large, with prominent tuberosity just posterior to orbital arch [S74] [A00]; postorbital forming lateral and anterior border of upper temporal fenestra [S74] [A00]; postorbital with short posterior process fitting under anterior process of squamosal [A00]; lower temporal fenestra approximately same size as orbit [S74]; squamosal with ventral process overlapping quadratojugal [A00]; squamosal with short, blunt posterior process [A00]; quadrate vertical with thick vertical column bearing anterior flange which overlaps quadratojugal [A00]; quadrate column curved posteriorly for to form border for tympanum [A00]; quadrate with convex articular condyle [A00]; supraoccipital contributes to foramen magnum [π$03]; supraoccipital triangular, fused to exoccipitals & opisthotics [A00]; exoccipitals fused to opisthotics and paroccipital process [A00]; paroccipital process not fully ossified distally [A00]; metotic foramen communicates nerves IX-XI, with separate hypoglossal foramen posterior to metotic foramen [A00]; paroccipital process forms base of posttemporal foramen [A00]; basioccipital forms most of occipital condyle [A00]; condyle bearing notochordal pit [A00]; basioccipital broadly overlaps basisphenoid [A00]; basioccipital with well-developed basal tubers [A00]; basioccipital with blind basioccipital recess [A00]; semilunar depression absent from braincase [G02]; basisphenoid dorsoventrally short [G02]; presphenoid may [A00] or may not [G02] be present above trigeminal notch; olfactory bulb at end of long peduncle [A00]; maxilla with palatal process massive, meeting on midline and vaulted, meeting vomer (also contact with palatine & ectopterygoid) [S74] [A00]; long triangular interpterygoid vacuity present [S74]; vomers not contacting premaxilla (but see figure) [A00]; palatines thin, with overlapping contacts [A00]; palatines, and palate generally, arches upward sharply [A00]; pterygoid large, contacting, vomer, ectopterygoid, palatine, quadrate & basisphenoid, bearing the usual palatal, transverse and quadrate rami [A00]; anterior ramus strongly convex ventrally & near vertical at midline [A00]; ectopterygoid with expanded lateral foot contacting jugal [A00]; dentary robust, not dorsoventrally expanded, & external surface is almost flat [A00]; dentary medial surface almost covered by splenial & coronoid [A00]; dentary symphysis with two flanges grasping opposite ramus [A00]; palatal dentition absent [S74] [P93]; teeth homodont, large, robust, laterally compressed, recurved, sharply pointed & serrated [S74]; tooth roots twice as long as crowns [S74]; teeth with thin, striated enamel [S74]; 4 teeth on premaxilla, 10 on maxilla [S74] (contra [A00] 12 on maxilla); interdental plates present [S74] [A00]; vertebrae amphicoelous [S74]; vertebrae with neural spines anteroposteriorly elongate & flat [S74]; vertebrae with prominent interspinous notch on anterior face [S74]; vertebrae with grossly constricted centra, leaving thin, vertical plate between flared rims [S74]; 23-25 presacral vertebrae [S74]; dorsal vertebrae compressed laterally & reduced to thin vertical plate between flared ends [S74]; rib facets well separated & on different levels throughout column, but never on centra [S74]; transverse processes short [S74]; prezygapophyses short, not overhanging centrum [S74]; neural spines flat, tall, thin, not inclined [S74]; ribs massive, with prominent ridge on proximal anterior surface [S74]; 2 sacral vertebrae, not fused & generally similar to dorsals [S74]; 2nd sacral & 1st caudal with anterior centrum flare enclosing and immobilizing end of preceding centrum [S74]; sacral transverse processes massive [S74]; caudal vertebrae shorter & with large rounded rims, less strongly constricted than dorsals (& 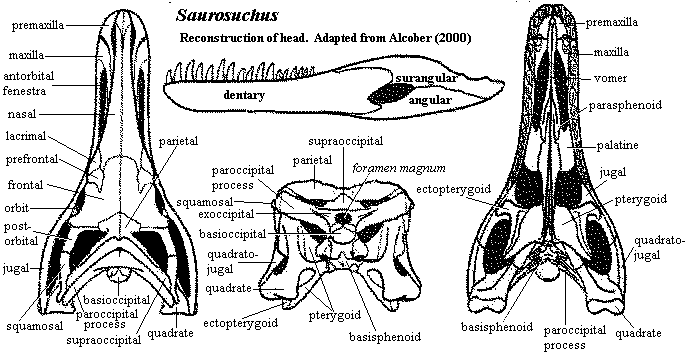 becoming even less constricted distally) [S74]; caudal prezygapophyses elongate distally, overhanging centra [S74]; shoulder girdle & forelimbs unknown [S74]; ilium with broad brevis shelf [S74]; ilium posterior blade long & heavy [S74]; ilium anterior blade short and emarginated ventrally [S74]; ischium long, rodlike, somewhat expanded distally & fused at midline over most of its length [S74]; ischium participates little in acetabulum [S74]; ischial blade dacriform in cross-section, with thin part forming symphysis [S74]; pubis almost excluded from acetabulum [S74]; pubis broad proximally, thinning rapidly to rod-like proportions [S74]; acetabulum high up on ilium, large, deep, & closed [S74]; femur with dacriform head [S74] femur slightly S-shaped [S74]; greater & lesser trochanters absent [S74]; 4th trochanter present as small rugose bulge [S74]; femur with long posterior flange over proximal 1/2 of length [S74]; tibia length 80% of femur, with cnemial crest extending 1/3 shaft length, separated by broad flat region from lateral condyle [S74]; tibia flares distally to broad articulating surface (as broad as distal femur) for astragalus & calcaneum [S74]; fibula with prominent iliofibularis tubercle [S74]; croc-normal tarsus [S74]; astragalus (anteroposteriorly?) shallow, with tibial trochlea & fibular articulations separated by a steep step, & bulbous anterior surface for metatarsal I [S74]; in articulation, astragalus & calcaneum line up so that fibular articulations of both are adjacent & form a sort of double ball & socket joint [S74]; calcaneum with posterior upturned "heel" [S74]; calcaneal tuber with lateral side concave & medial side convex [P93]; fourth tarsal large, tetrahedral [1], with prominent rounded facet for metatarsal V [S74]; only distal tarsals III & IV present [S74]; pes with 5 digits [S74]; metatarsal II largest [S74]; metatarsals II-IV of similar length, with III somewhat slender and IV concave laterally and closely appressed to III [2] [S74]; metatarsal V widely separated, hooked, & with broad medial expansion [S74]; pes 1&2 robust [S74]; pes 3 longest [S74]; pes 5 broad, flat & pointed laterally [S74]; phalangeal formula 2345(1-3) [S74]; dermal armor present as two paramedian rows of small, leaf-shaped, imbricating scutes, drawn to a point anteriorly, with these stems being overlapped by the next anterior scute [S74].
becoming even less constricted distally) [S74]; caudal prezygapophyses elongate distally, overhanging centra [S74]; shoulder girdle & forelimbs unknown [S74]; ilium with broad brevis shelf [S74]; ilium posterior blade long & heavy [S74]; ilium anterior blade short and emarginated ventrally [S74]; ischium long, rodlike, somewhat expanded distally & fused at midline over most of its length [S74]; ischium participates little in acetabulum [S74]; ischial blade dacriform in cross-section, with thin part forming symphysis [S74]; pubis almost excluded from acetabulum [S74]; pubis broad proximally, thinning rapidly to rod-like proportions [S74]; acetabulum high up on ilium, large, deep, & closed [S74]; femur with dacriform head [S74] femur slightly S-shaped [S74]; greater & lesser trochanters absent [S74]; 4th trochanter present as small rugose bulge [S74]; femur with long posterior flange over proximal 1/2 of length [S74]; tibia length 80% of femur, with cnemial crest extending 1/3 shaft length, separated by broad flat region from lateral condyle [S74]; tibia flares distally to broad articulating surface (as broad as distal femur) for astragalus & calcaneum [S74]; fibula with prominent iliofibularis tubercle [S74]; croc-normal tarsus [S74]; astragalus (anteroposteriorly?) shallow, with tibial trochlea & fibular articulations separated by a steep step, & bulbous anterior surface for metatarsal I [S74]; in articulation, astragalus & calcaneum line up so that fibular articulations of both are adjacent & form a sort of double ball & socket joint [S74]; calcaneum with posterior upturned "heel" [S74]; calcaneal tuber with lateral side concave & medial side convex [P93]; fourth tarsal large, tetrahedral [1], with prominent rounded facet for metatarsal V [S74]; only distal tarsals III & IV present [S74]; pes with 5 digits [S74]; metatarsal II largest [S74]; metatarsals II-IV of similar length, with III somewhat slender and IV concave laterally and closely appressed to III [2] [S74]; metatarsal V widely separated, hooked, & with broad medial expansion [S74]; pes 1&2 robust [S74]; pes 3 longest [S74]; pes 5 broad, flat & pointed laterally [S74]; phalangeal formula 2345(1-3) [S74]; dermal armor present as two paramedian rows of small, leaf-shaped, imbricating scutes, drawn to a point anteriorly, with these stems being overlapped by the next anterior scute [S74].
Notes: [1] Sill [S74] states at one point that distal tarsal IV is "subtriangular" [S74: 322]and elsewhere that it is "tetrahedral" [S74: 346]. His photograph suggests that the latter is correct. [2] That is, the metatarsals approach the arctometatarsalian condition of certain theropods. [3] "[π03]" refers to our own analysis of published data.
Links: UTDMG--Saurosuchus galilei.
References: Alcober (2000) [A00], Benton (1984) [B84], Bonaparte 1971) [B71], Gower (2002) [G02], Parrish (1993) [P93], Sill 1974) [S74]. MAK991019, ATW031128.
checked ATW060119
Using this material. All material by ATW is public domain and may be freely used in any way (also any material jointly written by ATW and MAK). All material by MAK is licensed Creative Commons Attribution License Version 3.0, and may be freely used provided acknowedgement is given. All Wikipedia material is either Gnu Open Source or Creative Commons (see original Wikipedia page for details). Other graphics are copyright their respective owners