Archosauria: Archosauria: Ornithodira
Scleromochlus
Taxa on This Page
- Crurotarsi
- Ornithodira
- Scleromochlus
|
|
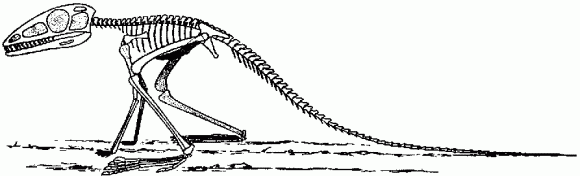
Scleromochlus taylori, a basal ornithodire from the Late Carnian Lossiemouth Beds of Elgin, Scotland. Reconstructed skeleton, length about 25 cm. Illustration in Krebs (1976) after Huene 1914). According to Benton 1999), Huene's reconstruction has the torso too short, the coracoid too small, and perhaps the feet a bit large.
|
The Stem Groups: Ornithodira and Crurotarsi
By definition, the Archosauria (rocs + crocs) are comprised of the Ornithodira (rocs > crocs) and the Crurotarsi crocs > rocs). The ornithodires went on to produce pterosaurs and dinosaurs, including the birds. Today, the Crurotarsi are represented only by the Crocodylia. However, the crurotarsans were responsible for most known reptile diversity in the Early and Middle Triassic. The only known basal ornithodire is the Scleromochlus from the Carnian of Scotland.
Scleromochlus and the Ornithodires
Ornithodires are supposed to share a number of characteristics, many of which are listed in the main entry below. Generally, ornithodires have moderately large heads with numerous fenestrae. In fact the ornithodire head often resembles a complex series of bony arches, rather than a box with holes. The neck has only 7 vertebrae -- sometimes a few more. The interclavicle is absent and the forelimbs are usually markedly shorter than the hindlimbs. The femur is bowed over most of its length, and the legs are generally located under the body. Reduction of the femoral fourth trochanter shows up as a synapomorphy in our analysis, which is surprising and possibly wrong. However, the structure of the limb bones is certainly simpler, with cleaner lines than in more basal archosauriforms. Likewise the ankle joint is strong, but simple and hinge-like. The foot is functionally tridactyl, with digits II - IV bound together and largely functioning as a single unit. The limb bones are quite often hollow, and there was primitively no body armor.
This body plan is one well-adapted for extended (or frequent) locomotion. It is very light, but strong. It is not particularly flexible. Rather, it's the sort of body an engineer might design to perform a series of kinetic and complex, but stereotyped, motions under a wide range of environmental conditions: tasks like running, jumping, or flying. But only one such task, and generally without much variation.
Scleromochlus makes an interesting example of that kind of body plan because it is not really very much like the only other two examples we have: dinosaurs and pterosaurs. Scleromochlus was small, about the size of a smallish cat, but much lighter. The head is somewhat like a pterosaur in general form but lacks any of the specializations of that clade. However, it is not merely a generalized ornithodire skull. Scleromochlus had its own suite of unique cranial specializations such as the broad nasal bones extending like an awning over the nostrils, and the conspicuously raised rim of the maxilla around the antorbital fossa. One might speculate that these anchored flaps of integument to protect exposed tissues from abrasion. That would be consistent with the low, triangular skull -- almost like a snake's head. However, the rest of the body looks like a leaping form: fairly short arms, long, simplified legs with a strong pelvis and heavily reinforced acetabulum, relatively compact torso and a long tail.
The object is not to reconstruct the lifestyle of Scleromochlus. Some of the very best minds of the last century have tried and failed to determine how these characteristics fit together. The lesson is rather that it is clearly an ornithodire, but is quite unlike either of the other ornithodire groups. The valid conclusion we may draw is that the Middle Triassic may well have contained a wealth of hidden ornithodire diversity. With three such disparate forms built on a body plan well-designed for occupying specialized niches, one suspects that the hot, dry inland areas of the Middle Triassic were home to any number of small, strange ornithodires, hopping, flapping, leaping and sprinting from one scarce resource to another.
Crurotarsi
The crurotarsans were made of sterner, if less athletic, stuff. Since the fossil record of basal Crurotarsi is reasonably good, we will not spend much time on a theoretical morphotype. They were generally large, and sometimes huge. The skull, although often strongly fenestrated, is usually far more massively constructed than the ornithodire norm. Basal Crurotarsi have a wide variety of specializations of the premaxilla. While this does not appear on anyone's list of synapomorphies, this feature is -- one might say -- as plain as the nose on their faces. The snout is typically very narrow and variably elongate, and the premaxilla is partially separated from the maxilla and/or forms a long anterior extension. This is difficult to pin down as a morphological character. However, it may well represent a developmental synapomorphy in which the premaxilla and maxilla develop quite separately, with less developmental interaction than is the case in most tetrapods. The cheek bones have a marked tendency to be laterally convex, often to the point of forming a sort of shelf around the side of the skull.
The crurotarsan neck is usually short and massive, with short, stout cervical ribs anchoring the musculature needed to move the massive head and neck. Not surprisingly, crurotarsans are much more likely to be obligate quadrupeds than are ornithodires. The limbs tend to be complex, with the tarsus and foot retaining many small bones and a fuller complement of functional digits. The limb posture may be sprawling or nearly vertical. However, Crurotarsi with vertically-oriented legs normally accomplish this posture by angling the acetabulum to face ventrally, rather than by bending the femur or developing a distinct femoral neck and head.
As noted on a previous page, crurotarsan phylogeny is poorly understood. Thus, it is hazardous to point to one feature or another as basal or derived except in the context of particular lineages. These will be surveyed on the following pages.
ATW040124
Descriptions
Ornithodira: Rocs > Crocs (not the same as Ornithodira sensu Benton = dinosaurs + pterosaurs)
Range: from the Middle Triassic.
Phylogeny: Archosauria: Crurotarsi+ * : Scleromochlus + (Pterosauria+ Dinosauromorpha).
Characters: Anterior cervical vertebrae longer than mid-dorsals [$S91]; "incipient" third sacral is either fully incorporated into sacrum or lost [$π03]; no interclavicle [$S91]; clavicles reduced or absent [$S91]; deltopectoral crest rectangular [S$91]; femoral shaft bowed anteriorly for at least 80% of length [$S91]; femoral 4th trochanter absent [$π03]; tibia > femur [S$91] [$π03]; astragalus without posterior groove [$S91]; astragalus & calcaneum fused to crus, mesotarsal ankle; calcaneal tuber small or absent [$S91]; calcaneal tuber does not flare reversal) [P93]; distal tarsals 3 & 4 of similar width [$S91] [$π03]; metatarsals bunched [$S91] [$π03]; metatarsals II-IV elongate; metatarsals II-IV longer than 50% tibia length [$S91] [$π03]; digitigrade on pes II-IV; dorsal body osteoderms absent [S$91][$π03].
Notes: [1] "[π03]" refers to our own analysis of published data. [2] The image seems to be based in part on Huene's reconstruction which underestimates the length of the dorsal vertebral column [B99].
Links: link
References: Benton 1999) [B99]; Parrish 1993) [P93], Sereno (1991) [S91], Unwin 1999). ATW040118.
 Scleromochlus: S. taylori Woodward, 1907 [B99].
Scleromochlus: S. taylori Woodward, 1907 [B99].
Range: Late Triassic (Carnian) of Europe (Scotland).
Phylogeny: Ornithodira: Pterosauria + Dinosauromorpha) + *.
Characters: length ~18cm [B99] [3]; skull low & triangular, twice as broad across orbits as deep [$S91] [B99]; skull more than 50% length of presacral vertebral column [$π03] [2]; premaxilla elongate [B99]; maxilla with narrow process to nasal anterior to antorbital fossa & forming most of interior of fossa [B99]; maxilla forming raised anterior margin of antorbital fossa [$S91]; maxilla meets lacrimal - prefrontal between orbit & antorbital fossa & meets jugal posteriorly [B99]; expanded nasal forms anterior plate hiding nares & premaxilla in dorsal view [B99]; nasal descending flange forms dorsal wall of antorbital fossa [B99]; frontals form most of dorsal orbit [B99]; parietals broad, expanding behind orbits, with straight posterior margin [B99]; small postfrontal may be present [B99]; UTF present as a narrow slit between parietal and postorbital - squamosal [B99]; lacrimal & prefrontal fused, forming bar between orbit & antorbital fossa [B99]; orbit large & nearly circular [B99]; jugal with broad dorsal process meeting postorbital behind orbit and narrow posterior process meeting quadratojugal in long, diagonal suture [B99]; quadratojugal with quadrate facet well forward, at about level of basipterygoid process [B99]; braincase long [B99]; basipterygoid processes long & massive [B99]; sella turcica narrow [B99]; premaxilla forming short platform on anterior palate [B99]; palatal elements as shown in image [B99]; palatine with plate-like process meeting maxilla [B99] [4]; long interpterygoid vacuity [B99]; pterygoids with deep recess for basipterygoid process [B99]; lower jaw slender, not straight in lateral view [B99]; dentary symphysis short & strong, wedge-shaped in lateral view [B99]; long retroarticular process bent medially [B99]; teeth isodont & very small [B99]; premaxilla with ~6 teeth [B99]; maxilla with ~10 teeth (distal 1/3 edentulous), dentary with ~15 [B99]; presacral vertebrae very short [W70]; at least 8 cervicals & perhaps 16-17 dorsals [B99]; cervicals slightly shorter than dorsals [B99]; centra spool-shaped, amphicoelous with deep central depressions & constricted laterally & ventrally [B99]; short cervical ribs present [B99]; neural arches long & low in cervicals, taller & with long transverse processes in dorsals [B99]; gastralia present [B99]; 3-4 sacral vertebrae [B99] [$π03]; about 35 caudals [B99]; caudals with posteriorly slanting neural spines [B99]; limb bones hollow [B99];scapula long & slender, tapering distally [B99]; coracoid not elongate, but short & oval [W70] [B99]; forelimb <55% 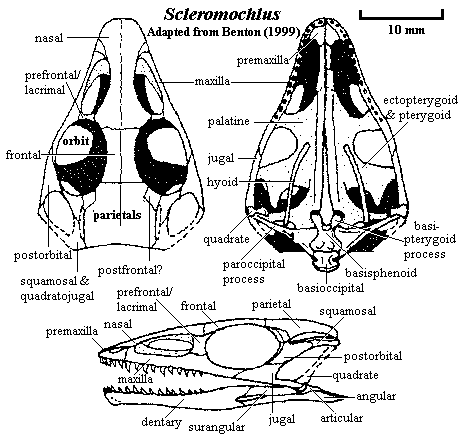 length of hindlimb [$π03]; humerus/femur ratio 0.61 [W70]; humerus long & thin, with short, well-defined deltopectoral crest, deeply divided on ventral face distally [B99]; radius & ulna only slightly shorter than humerus [W70] [B99]; carpus & manus short, with 5 metacarpals of similar length (McI may be shorter) [B99]; pelvis small, with low iliac blade & short pubis & ischium [B99]; strong supraacetabular crest present [B99] [$π03]; iliac blades elongate [B99]; pubis longer than ischium [B99] [$π03]; pubis with heavy acetabular margin and possible ventromedial process [B99]; ischial foot absent [B99]; acetabulum closed [B99]; femur slender & without trochanters, except a possible low 4th trochanter [B99]; proximal femur expanded longitudinally & distal femur expanded laterally [B99]; tibia longer than femur [W70] [1]; tibia distally and proximally) expanded, with rectangular end [B99] [$π03]; astragalus not fused to tibia & without ascending process [B99]; [5]; tarsus crocodile-normal [W70] (contra [B99]: astragalus & calcaneum of similar size, and not clear which is which, thus ankle structure is uncertain); metatarsals I-IV roughly equal in length [W70] and closely appressed [B99]; metatarsals II-IV longer than 50% of tibia length [B99] [$π03]; MtV short & probably without phalanges [B99]; cranial bones without ornament [B99]; very thin scutes or keratinous scales probably present [B99].
length of hindlimb [$π03]; humerus/femur ratio 0.61 [W70]; humerus long & thin, with short, well-defined deltopectoral crest, deeply divided on ventral face distally [B99]; radius & ulna only slightly shorter than humerus [W70] [B99]; carpus & manus short, with 5 metacarpals of similar length (McI may be shorter) [B99]; pelvis small, with low iliac blade & short pubis & ischium [B99]; strong supraacetabular crest present [B99] [$π03]; iliac blades elongate [B99]; pubis longer than ischium [B99] [$π03]; pubis with heavy acetabular margin and possible ventromedial process [B99]; ischial foot absent [B99]; acetabulum closed [B99]; femur slender & without trochanters, except a possible low 4th trochanter [B99]; proximal femur expanded longitudinally & distal femur expanded laterally [B99]; tibia longer than femur [W70] [1]; tibia distally and proximally) expanded, with rectangular end [B99] [$π03]; astragalus not fused to tibia & without ascending process [B99]; [5]; tarsus crocodile-normal [W70] (contra [B99]: astragalus & calcaneum of similar size, and not clear which is which, thus ankle structure is uncertain); metatarsals I-IV roughly equal in length [W70] and closely appressed [B99]; metatarsals II-IV longer than 50% of tibia length [B99] [$π03]; MtV short & probably without phalanges [B99]; cranial bones without ornament [B99]; very thin scutes or keratinous scales probably present [B99].
Notes: [1] Walker [W70: 361] states that the limb segment ratios "indicate a definite tendency towards bipedal hopping." Benton [B99] generally agrees, pointing to the elongation of the distal hindlimb, elongate tail, short anterior trunk, small pelvic girdle [B99]. However he notes that the se features are not present to quite the extent one would expect based on living mammal saltators. [2] "[π03]" refers to our own analysis of published data. [3] Interestingly, almost all of the 7 specimens were found in ?dorsal roadkill posture [B99], a fact with implications for its mode of life. [4] This is not clear in the images provided of the specimens themselves. [5] Benton [B99] states both that the fibula is "circular in cross-section" and that it is "flattened." [B99: 1432]. Apparently it is flattened against the tibia only for a small distance in part of the proximal half.
Image: adapted from a drawing by John Sibbick in [B99].
Links: Scleromochlus -- The Dinosauricon some nice images); 恐竜データ集 -ス- Japanese); 川崎悟司イラスト集・スクレロモクルス Japanese).
References: Benton 1999) [B99]; Sereno (1991) [S91], Walker (1970) [W70].
 Crurotarsi: Crocs > Rocs
Crurotarsi: Crocs > Rocs
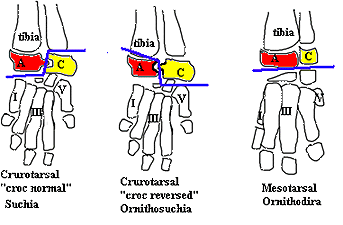
Characters: Cervical ribs short & stout [$π03]; scapulocoracoid with anterior notch between scapula & coracoid [$π03]; humerus with relatively distinct internal tuberosity, just distal to which the shaft is strongly curved [$S91]; fibula with anterior trochanter robust & knob-like [$S91]; fibula with distal end wider than proximal end [$S91]; crocodylomorph tarsus in which astragalus acts as extension of the crus & calcaneum associated with pes, with joint along articulation between astragalus & calcaneum [$P93]; ventral astragalocalcaneal facet small [$S91] [$π03]; astragalus with tibial facet flexed or convex [$S91] [$π03]; extended wheel-like ("hemicylindrical") process on calcaneum rotates fibula [$S91] [P93] [$π03]; articulated by peg in astragalus fitting into socket in calcaneum (croc normal) or the reverse croc-reverse); calcaneal tuber broader than long [$S91] [$π03]; calcaneal tuber with rounded ("flared" [S91]) end [$S91] [$π03]; fifth metatarsal remains large primitively and may have shared tarsal role; pes 5 with 4 phalanges [$π03]; single paramedian osteoderm pair per presacral vertebra [$S91].
Notes: "[π03]" refers to our own analysis of published data.
Links: Crurotarsi; Lecture 10 - Triassic: Newark, Chinle; Archosauria: More on Morphology; Pseudosuchia; Biology 356.
References: Parrish (1993) [P93], Sereno (1991) [S91]. ATW040103.
checked ATW050702
Using this material. All material by ATW is public domain and may be freely used in any way (also any material jointly written by ATW and MAK). All material by MAK is licensed Creative Commons Attribution License Version 3.0, and may be freely used provided acknowedgement is given. All Wikipedia material is either Gnu Open Source or Creative Commons (see original Wikipedia page for details). Other graphics are copyright their respective owners