Crocodylomorpha
Taxa on This Page
- Crocodylomorpha
- Sphenosuchia
- Sphenosuchidae
- Trialestidae
The Heart of the Crocodile
This essay replaces a small, cryptic paragraph previously included as "Crocodylia Notes." Like this effort, the previous note dealt with circulation. In fact, it is customary to accompany any discussion of crocodylians with a discussion of circulation for the simple reason crocs have a better, more flexible and more efficient heart than any other organism, alive or (presumably) dead. The croc system is also rather complex. Perhaps the idea is that if one can understand the croc heart, then anything else is a piece of cake.
Of course, more sinister motives may be at work. Students are most commonly introduced to croc circulation with a semi-schematic diagram which demonstrates what is surely a gleefully malicious intent to confuse. The left atrium and left ventricle are drawn on the right. This would be correct in ventral view, but the atria are on the bottom, which is contrary to the usual "valentine" representation of the heart, and also just plain wrong. Naturally, the left aorta emerges from the right ventricle, and vice-versa, while a connection between the aortas, the foramen of Panizza, is explained as creating a "right-to-left intercardiac shunt" when blood flows from the left aorta to the right aorta outside the heart. No reasonable student can view this representation without the deep suspicion that she is the butt of some bizarre, two-century-old practical joke handed down in the professoriate from one generation of cackling, gummy-eyed, misanthropic curmudgeons to the next.
Sadly, we are now probably doomed by nomenclature and convention endlessly to repeat this cycle of obfuscation, like a hideous parody of the circulatory system itself. However, in spite of the awful weight of history, an attempt will be made here, by casting off the shackles of nomenclature and concentrating on fundamentals. Nomenclature will, of necessity, be introduced as required, but not before.
The tetrapod circulatory system is essentially two circuits running off a single pump (Fig. 1). Blood alternates between picking up oxygen in the lung circuit and dropping it off in the body circuit. Because two circuits are involved, tetrapods functionally divide the heart into a left side (the body circuit) and a right side (the lung circuit). This functional division is manifested as a physical division in birds, mammals and crocs. However, turtles, lizards, and presumably pareiasaurs, mosasaurs, and so on achieve the same effect using partial separations and some rather neat hydrodynamic controls.
Each side of the heart is also divided, roughly, into an "in box" (atrium) and an "out box" (ventricle). In the conventional "valentine" representation of the heart, it is the atria which form the two upper lobes. With that symbol in mind, the figures here show the atria above the ventricles. In reality, the geometry of the heart is much more complex and somewhat variable.
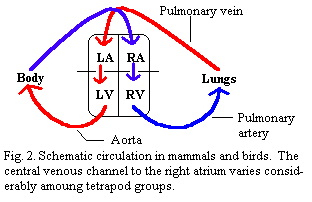 Birds and mammals have a relatively simple two compartment design. (Fig. 2) The lung and body compartments of the heart are not interconnected at all. Each blood cell must make the rounds of both circuits sequentially. One aorta emerges from the left ventricle and pumps blood to the body, which eventually returns, spent and hung over, at the right atrium. The pulmonary artery takes this blood from the right ventricle into the lungs and returns it refreshed and oxygenated, to the left atrium. That is, essentially, all that birds and mammals can manage.
Birds and mammals have a relatively simple two compartment design. (Fig. 2) The lung and body compartments of the heart are not interconnected at all. Each blood cell must make the rounds of both circuits sequentially. One aorta emerges from the left ventricle and pumps blood to the body, which eventually returns, spent and hung over, at the right atrium. The pulmonary artery takes this blood from the right ventricle into the lungs and returns it refreshed and oxygenated, to the left atrium. That is, essentially, all that birds and mammals can manage.
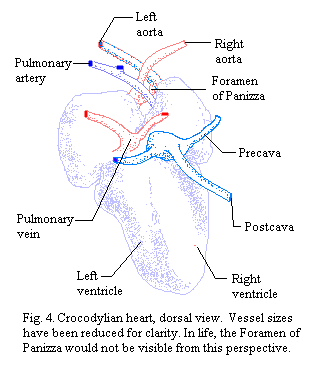
Crocs have two aortas, one starting from the left side and one from the right. Unfortunately, the aorta leading from the left side bends to the right, and vice-versa. Thus the aorta connected to the left side of the heart is the "right" aorta. The aorta from the right side is the "left" aorta. The right aorta passes blood more directly to the brain and anterior circulation, although it also directs some to more visceral and posterior regions. The flow from the left aorta is directed more exclusively to the viscera and posterior circulation. However, the two aortas are connected at the base by a shunt, the Foramen of Panizza. 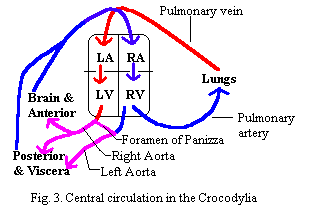 Fig. 3. As a result, some rather complex patterns of regulation become possible. For reference, Fig. 4 is included to illustrate a somewhat more realistic picture of the Crocodylian heart in dorsal view. Some of the vessels shown in this figure are discussed in more detail below.
Fig. 3. As a result, some rather complex patterns of regulation become possible. For reference, Fig. 4 is included to illustrate a somewhat more realistic picture of the Crocodylian heart in dorsal view. Some of the vessels shown in this figure are discussed in more detail below.
When blood pressure in the left and right compartments is equal, as in a croc at rest on land, there is little movement through the shunt. Oxygenated blood is preferentially received by the brain. In addition, more acidic, deoxygenated blood is preferentially received by the viscera where it is apparently used to produce digestive acids.
When the animal is active on land, circulation approaches the mammalian or avian condition. Blood pressure is high in the left compartment and oxygenated blood is shunted to the left aorta. Deoxygenated blood in the right compartment is almost entirely routed to the lungs.
Finally, when the croc dives, the pulmonary system is partially shut down, causing higher pressure on the right than on the left. As a consequence blood from the right ventricle flows into the right aorta. Since the animal is not breathing, there is no fresh oxygenated blood. Thus the full ventricular output of the heart can be used to circulate nutrients and filter waste products -- particularly important functions when oxidative metabolism cannot be used to break down metabolic wastes. The increased flow through the Left aorta also tends to retain body heat in the central, visceral circulation.
 These are the conventional explanations for the probably circulatory states of the croc heart. As mammals, we are accustomed to think of blood primarily as an oxygen transport mechanism. Our metabolic needs for oxygen are very high, and this perception is not really unfair. However, in vertebrates other than mammals, birds and perhaps other dinosaurs, the respiratory functions of circulation are not as predominant. This requires a more balanced and complex regimen of circulatory regulation, as seen in the Crocodylia. Unfortunately, studies of croc circulatory regulation seem to have focused largely on respiration. As a result, we may be partially missing the point. The rationale and mechanisms of this regulation may actually lie in waste clearance and transport, immunology, regeneration, pH and ion balances, metabolic signaling, and all of the other functions of circulation. --ATW
000521
These are the conventional explanations for the probably circulatory states of the croc heart. As mammals, we are accustomed to think of blood primarily as an oxygen transport mechanism. Our metabolic needs for oxygen are very high, and this perception is not really unfair. However, in vertebrates other than mammals, birds and perhaps other dinosaurs, the respiratory functions of circulation are not as predominant. This requires a more balanced and complex regimen of circulatory regulation, as seen in the Crocodylia. Unfortunately, studies of croc circulatory regulation seem to have focused largely on respiration. As a result, we may be partially missing the point. The rationale and mechanisms of this regulation may actually lie in waste clearance and transport, immunology, regeneration, pH and ion balances, metabolic signaling, and all of the other functions of circulation. --ATW
000521
The basic vertebrate arterial system is based on a one compartment design. That is, the heart pushes blood through the gill arches, through the various organs, and back to the heart. Primitively, then, there was a one-to-one correspondence between aortic arches and gill arches. In fact, one of the more convincing demonstrations of the monophyly of the gnathostomes is that all vertebrates with jaws begin the development of the arterial system with six pairs of aortic and gill arches. The number in jawless vertebrates is much more diverse and is normally larger (eight pairs in lampreys, perhaps ten in ostracoderms). In truth, this evolutionary and embryological unity has a great deal of aesthetic appeal. However, in highly derived forms such as crocs, this basal architecture has been largely replaced with an irregular, asymmetrical flow pattern and these primordial symmetries have relatively little functional significance.
 It may be more useful to think of the circulatory system in terms of its functional units. Every vertebrate circulatory system must accomplish at least the following two functions: gas transport and nutrient transport. For the (at least) two compartment tetrapod circulation, this may be broken down into a series of major sub-tasks as follows:
It may be more useful to think of the circulatory system in terms of its functional units. Every vertebrate circulatory system must accomplish at least the following two functions: gas transport and nutrient transport. For the (at least) two compartment tetrapod circulation, this may be broken down into a series of major sub-tasks as follows:
Gas Transport
1. carbon dioxide and oxygen transport between the heart and respiratory surfaces (usually the lungs);
2. gas transport and exchange with the major, mostly posterior, muscles of the body wall and limbs; and
3. gas transport and exchange with major anterior organs such as the brain and jaw.
Nutrient and Waste Transport
4. mobilization of nutrients from the digestive organs;
5. detoxification of digestive products in the liver; and
6. removal of soluble wastes in the kidneys.
In addition, of course, the circulatory system has critical functions related to ion and water balance, transport and delivery of hormones and other chemical signals, and the efficient delivery of life-saving quantities of alcohol to grateful neural receptors. However, the foregoing six functions are sufficient to explain the main patterns of circulation for the present purposes. Following the functional outline above (see also figures 1& 2):
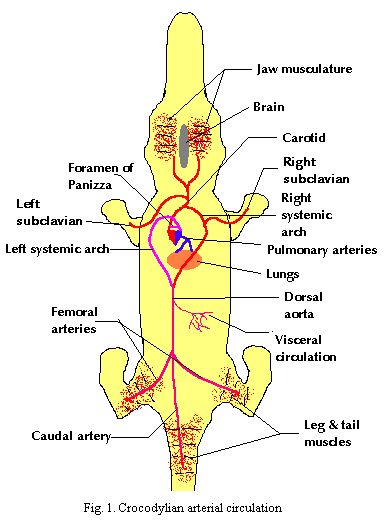
1. Gas transport in the heart-lung circuit is accomplished by the pulmonary aorta and pulmonary veins.
2. Gas transport to the posterior consumers occurs through both the left and right systemic arches as outlined in the prior essay. The left aorta joins with the posterior branch of the right aorta to form the dorsal aorta which curves posteriorly and trifurcates into the two femoral arteries and the caudal artery. Return circulation is through the caudal and iliac veins and either through the renal portal or through the lateral abdominal veins to the liver. In either case, the venous blood eventually reenters the heart through the postcava.
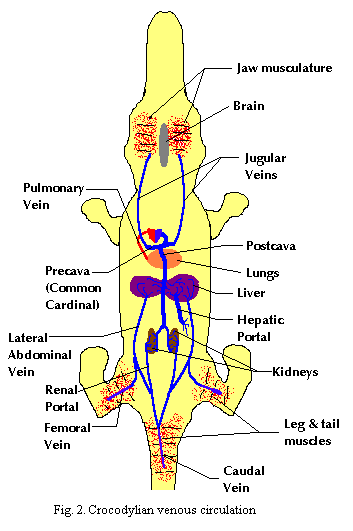 3. Gas transport to anterior consumers occurs exclusively through the left systemic arch, which gives rise to the carotid arteries. Return circulation proceeds through the jugulars into the precava or common cardinal vein.*
3. Gas transport to anterior consumers occurs exclusively through the left systemic arch, which gives rise to the carotid arteries. Return circulation proceeds through the jugulars into the precava or common cardinal vein.*
4&5. Circulation to the digestive organs also occurs through the dorsal aorta. Dissolved nutrients are returned through the hepatic portal, across the hepatic filtration system, and into the postcava.
6. As noted in (2), posterior circulation may pass through the renal portal vein, across the kidneys and, after renal filtration, to the postcava. ATW 000603.
Descriptions
Crocodylomorpha:
Skull massive and well-buttressed; no skull kinesis (skull absorbs force of strong, rapid bite); antorbital fenestrae small (closed in modern forms); descending process of prefrontal present [π$03]; postfrontals absent; descending process of squamosal absent [$P93] [π$03]; squamosal overhangs temporal region [π$03]; anterior extension of quadrate; quadrate complex; quadrate contacts prootic [π$03]; occiput solid, no or minor occipital fossae [π$03]; exoccipital with lateral crest lying anterior to hypoglossal foramen [$G02]; palate & braincase strongly integrated; ventral ramus of opisthotic makes extensive contact with prootic anteroventral to fenestra ovalis [π$03]; middle ear elements pneumatized [π$03]; crista vestibuli present [π$03]; lagenar recess visible externally as prominence on wall of otic capsule [π$03]; Eustachian tubes at least partially encased in bone [π$03]; prootic not in broad contact with anterior of paroccipital process [π$03]; external foramen for abducens nerve solely within basisphenoid [π$03]; epipterygoid absent; basipterygoid processes enlarged [π$03]; pila antotica ossified by basisphenoid and laterosphenoid so that these elements are in contact anterior to trigeminal foramen [π$03]; secondary palate formed by maxillae & vomers [$P93]; large anterior & posterior pterygoideus muscles, oriented horizontally; parasphenoid rostrum forms vertically expanded wedge [π$03]; skull bones pneumatized; 24 presacral vertebrae, 2 sacral vertebrae; accessory neural spine on mid-caudal vertebrae [π$03]; cartilaginous sternum (modern forms) separates posteroventral extensions of coracoids; no clavicle (all suggest obligate quadruped); elongated carpals (ulnare and radiale) perhaps involved in giving extra swing to foreleg [$P93]; manus IV with 5 phalanges (reversal) [π$03]; pubic shaft length >3x width of acetabulum [π$03]; pubis excluded from acetabulum since Late Jurassic; acetabulum semi-perforate [π$03]; femoral head without distinct neck, but bent medially with parallel medial & lateral sides [$P93]; metatarsal I length at least 85% length of Mt III [π$03]; pes IV with <5 phalanges [$P93]; pes V with 2 phalanges [π$03]; paramedial osteoderms sculptured [$P93]; osteoderms with anterolateral articulatory process & dorsolateral keel (as in Euscolosuchus) [S92].
Notes: [1] "[π03]" refers to our own analysis of published data.
Links: The Forgotten Archosaurs - infpormative blog dedicated to fossil crocs, gwu faculty clark image (cladogram); columbia; Crocodylomorpha; Crocodylomorpha -- The Dinosauricon; Reptilia.html; Basal Crocodylomorpha [Sphenosuchia] after Clark, Sues & Berman, ... Mikko's Phylogeny); Crocodylomorpha basic systematics, with some references); CROC LINKS some individual links and pointers to the Big Three croc sites); Those three sites are: Crocodilians: Natural History & Conservation: Crocodiles, Caimans, Alligators, Gharials Crocodilian.com), The Bibliography of Crocodilian Biology still extraordinary bibliograhical reference from Prof. Mason Meers, though now a bit dated), and Crocodile Specialist Group mostly a conservation site); RAPID COMMUNICATION FIRST RECORD OF ERPETOSUCHUS (REPTILIA- .... one of two relatively recent papers discussing Erpetosuchus -- the sister of Crocodylomorpha); Steve Turner's Crocodilian Link Page another good links page, but also somewhat dated).
References: Gower (2002) [G02], Parrish (1993) [P93], Sues (1992) [S92]. ATW031223.
Sphenosuchia: Gracilisuchus?
(like stem Archosauriforms), Sphenosuchus.
Range: Middle to Late Triassic
Apparently characterized by absence of derived characters(?!). Pneumatization limited; posterior process of prefrontal turns onto orbital surface of frontal where it inserts into a groove, rather than maintaining its usual superficial position on the dorsal skull roof; rim on lateral margin of supratemporal fossa; squamosal arcuate, flaring, with low-angle articular surface for the postorbital; squamosal with forked posterior process, straddling paroccipital process; quadrate does not contact lateral wall of braincase; little or no secondary palate.
Links: link; crocodylomorpha.
Trialestidae: Trialestes(= "Triassolestes")
Range: Late Triassic of South America.
Phylogeny: Sphenosuchia : Sphenosuchidae + *.
Characters: small, lightly built animal; elongate limbs; fore and hind legs of equal size; elongate carpals [C+00]; digitigrade; well-developed supraacetabular crest (like dinosaurs) [C+00]; acetabulum perforate (like dinosaurs) [C+00]; medium-sized terrestrial insectivore/carnivore.
Note: may turn out to be a primitive dinosaur. The again, it may be a crocodylomorph, but not a sphenosuchian. The third possibility is that specimens from different taxa have been referred to this single species. See [C+00].
Links: Trialestes; New References; Trialestes.
References: Clark et al. 2000) [C+00]. ATW030625
Sphenosuchidae: Dibothrosuchus Early Jurassic of China), Pedeticosaurus (Early Jurassic of South Africa), Platyognathus Early Jurassic of China), Sphenosuchus (Early Jurassic of South Africa), Terrestrisuchus (Late Triassic of England).
Range: Middle Triassic to Early Jurassic of Europe, North America, South Africa and China(?).
Phylogeny: Sphenosuchia : Trialestidae + *.
Characters: Loss of postfrontal unites sphenosuchids to crocodiles; erect, bipedal posture; terrestrial.
Links: All About Archaeopteryx (historical interest); Biology 356; Nathis Fauna Krokodillen (Dutch). 020317.
{kind=link}