 Faster then a speeding bullet?
Faster then a speeding bullet?| Coelurosauria | ||
| The Vertebrates | Tyrannosauroidea |
| Vertebrates Home | Vertebrate | Vertebrate |
|
Abbreviated Dendrogram
Dinosauria
├─Ornithischia
└─┬─Sauropodomorpha
└─Theropoda
│
└─Coelurosauria
├─Tyrannosauroidea
│ ├─Eotyrannus
│ └─Tyrannosauridae
└─Maniraptora
├─Therizinosauroidea
└─┬─Alvarezsauridae
└─┬─Oviraptorosauria
└─┬─┬─Troodontidae
│ └─Dromaeosauridae
└─Aves
|
Contents
Index |
Tyrannosaurs have generated a remarkable number of interesting disputes over the years. It would be impossible for us to add anything of merit to the various debates -- which, of course, will not discourage us from dispensing opinionated drivel as we see fit. Most of the issues are spin-offs from the hunter-scavenger debate: were the tyrannosaurs superpredators, gigantic garbage disposals, or something else? Saying anything about the behavior of long-extinct animals is chancey. Hypotheses in this area tend to be inherently unscientific because they are difficult to test rigorously. We are forced to measure the unknowable indirectly, using criteria that are themselves uncertain. So, were the tyrannosaurs "superpredators"? Fortunately, when it comes to "superness," there are certain widely accepted tests which we may apply.
Faster then a speeding bullet?One of the more astounding claims made for Tyrannosaurus is that it was able to run at 20m/sec (72 kph) or more. See, e.g., Bakker 1986) and several papers by Gregory Paul. [1]. However, the supposition is not as unreasonable as it may seem at first. Tyrannosaurus was very lightly built for its size, with long legs and enormous potential for muscle development in the right places. Its legs seem at least arguably capable of standing the shock of repeated impacts, and its excellent vision might allow it to navigate at speed. We cannot dismiss the possibility that it was capable of near-highway speeds for at least short sprints.
Nevertheless, we tend to doubt that tyrannosaurs were capable of quite such athleticism. Two critical studies on the subject have taken different approaches, but have arrived at similar negative conclusions. The first is by one of the masters of Mesozoic mechanics, John Hutchinson. Hutchinson & Garcia (2002). Hutchinson probably knows as much as anyone about theropod hindlimb muscles and locomotion. See Hutchinson 2001), Hutchinson 2001a) and Hutchinson & Gatesy (2000). He has not only rewritten the book on theropod myology, but has spent a good many years applying it to locomotion studies. Hutchinson & Garcia take advantage of this detailed knowledge of musculature to calculate the minimum percentage of body mass which must be devoted to leg extensors in order to keep a running tyrannosaur moving.
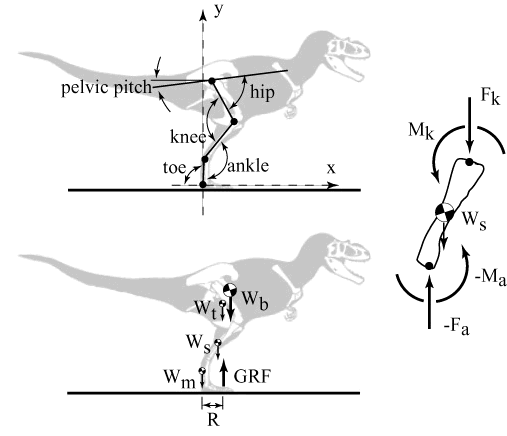
Hutchinson & Garcia apply the simplifying assumptions available at mid-stance phase (shown in the figure). This is the point during the running cycle when the body's center of gravity is lowest. At this stage of the running cycle, one foot is squarely on the ground, the vertical velocity is zero, and all of the force is directed straight up (d2x/dt2 = dy/dt = 0). Now, forget about running for the moment and assume that the dinosaur is just standing on one leg. Yes, he's off balance, but we're only interested in forces for the moment, and all the forces are vertical at this point in the running cycle. Since, for this fraction of a second, the body is not moving vertically, we know that the force of the animal against the ground is equal to its weight (W), and that the equal and opposite ground reaction force (GRF) will be the same. Newton's First Law, right? Now look at the diagram of the lower leg over on the right of the image. -Fa is the force at the ankle and Fk is the weight of the animal above the knee. These numbers aren't quite the same as W and GRF because we have to adjust for the weight of the foot below the lower leg and the weight of the shank itself. However, those are easy to estimate. The point is that two vertical forces, both of which we can calculate, are acting at opposite ends of the shank. Since the lower leg is not vertical, these forces tend to rotate it as shown. We can make good, conservative estimates of the angles involved. [2] Thus, we can calculate the torque as a function of the animal's weight. If the tyrannosaur has zero vertical velocity, we can then calculate the force needed to keep the leg from collapsing. Since we know, generally, where the muscles are, how strong vertebrate muscle is, where the muscles attach, and the angles these attachments make to the bone, we can estimate the amount of muscle needed to generate this force -- again as a function of the weight of the tyrannosaur. Finally, because this is a running dinosaur, we apply a "fudge factor" to allow  for the fact that the beast has to be accelerating vertically to make his next step. This turns out to be a fairly constant number (2.5) in all vertebrates. Obviously, the number gets higher for faster running speeds, but 2.5 does it for the basic running cycle. Actually, H&G's calculation is much more complex, since they used a model with at least 2 segments which could rotate independently. However, the basic approach is as described.)
for the fact that the beast has to be accelerating vertically to make his next step. This turns out to be a fairly constant number (2.5) in all vertebrates. Obviously, the number gets higher for faster running speeds, but 2.5 does it for the basic running cycle. Actually, H&G's calculation is much more complex, since they used a model with at least 2 segments which could rotate independently. However, the basic approach is as described.)
Because we have calculated everything as a function of weight, we don't have to know how heavy our tyrannosaur is. The answer is expressed in terms of muscle mass as a percentage of total mass of the dinosaur. Hutchinson & Garcia tested their model on a number of systems: people, chickens and alligators. These gave good results and indicated that bipedal runners usually have about twice the muscle mass they theoretically "need" as a safety factor. (In fact, safety factors of 2-3 seem to be an evolutionary constant). However, even without a safety factor, a running Tyrannosaurus would need to devote about 83% of its mass to leg extensors, which is plainly impossible.
As Hutchinson & Garcia point out, this doesn't really relate to the scavenger/predator issue because the same general parameters would apply to tyrannosaur prey animals. However, a similar result was reached by Farlow et al. (1995), using a completely different approach which is somewhat more specific to large, obligate bipeds like the tyrannosauroids. In discussing this paper, we will make the assumption that the reader has had about all the math she wants to deal with in the preceding paragraphs. Accordingly, we will deal only in qualitative terms. In essence, Farlow & Co. ask what would happen if a roughly 6000 kg Tyrannosaurus, moving at 20 m/sec, stepped on the Mesozoic equivalent of a banana peel. This is a fair question. As one might expect, the results of a 6 ton object, moving at 70 kph, hitting the ground from a height of a couple of meters are not pretty and would very likely be lethal for the object involved. Farlow et al. make the calculations based several different assumptions about substrate and come up with a train wreck each time. These results are not due to pessimistic assumptions. In fact the authors ignore effects which would make the injuries considerably more serious. [3]. Such accidents go to the heart of the evolutionary process. Species which engage in risky behaviors do not survive to reproduce unless the rewards are at least high enough to justify the risk.
 It's a Bird?
It's a Bird?Definitely not. The teeth of tyrannosauroids, particularly Tyrannosaurus, are unusual among theropods because they are individually large and heavy. There is little question that these teeth are capable of crushing bone. See image under Tyrannosaurinae. The question is whether tyrannosauroids regularly did so and under what circumstances. The assumption in the literature seems to be that bone-crushing is correlated with a scavenging life-style. Thus, those who view tyrannosaurs as cowardly scavengers tend to treat this feature as a point in their favor. Those who view tyrannosauroids as noble hunters tend to minimize the significance of this behavior.
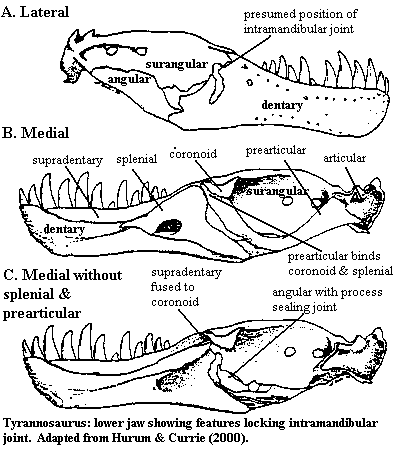
One factor which seems to favor bone-crushing as a significant behavior is the design of the tyrannosauroid jaw. Older reconstruction of tyrannosaurs usually incorporated the typical theropod jaw, which includes an intramandibular joint. This joint connects the anterior dentary, splenial and (if present) supradentary with the posterior surangular, angular, coronoid, prearticular and articular. Since this connection is hinged, the lower jaw bends outward in the middle when it is stressed -- as, for example, when the teeth hit something rather hard. Accordingly, most bone would not be crushed. Rather the teeth would slide over it as the jaw deformed, causing the bone to be swallowed whole or rejected.
This arrangement is certainly operative in carnosaurs, and perhaps even basal tyrannosauroids. However, Hurum & Currie (2000) have shown that tyrannosaurinids block the joint. The supradentary overgrows the joint and fuses with the coronoid. This connection is reinforced by a long process of the prearticular which articulates with both the coronoid and the splenial. An anterior process of the angular also bridges the gap ventrally. In addition, the supradentary sends ridges between the teeth, further immobilizing the lower tooth row.
Farlow & Holtz (2002) state that "there is no indication that tyrannosaurids deliberately crushed bones, in the manner of some mammalian carnivores, even though their teeth and jaws were strong enough to handle bone-breaking ... bone-biting seems to have been incidental to feeding on meat." The problem here is the "deliberate" part. Unlike first degree murder, bone-crushing requires no legal scientur. Mens rea, or an evil state of mind, is not an element of the offense to be proven. Tyrannosaurs seem to have evolved in a direction from which we may infer selective pressure to treat bone as edible.
Buchholtz (1997) notes that broken teeth are extremely common among tyrannosauroids. However this is not a universal trait. Lambe 1917) remarks, with some surprise, that the teeth of his Gorgosaurus showed no signs of wear at all. Fortunately we have at least two direct indications of tyrannosauroid diets: one enormous coprolite and some fossilized stomach contents. Varricchio 2001). Both seem to contain an unreasonable amount of juvenile hadrosaur bone. On the basis of this completely inadequate sample, we may speculate that, for the type of prey normally consumed by tyrannosaurs, their dentition and jaw mechanics simply allowed them to treat bone as irrelevant.
Farlow & Holtz (2002) discuss the work of Farlow on range sizes for large predators. Range sizes for living predators are closely correlated with body mass. If that relationship applies to Mesozoic predators, the home range size of the 2500 kg Acrocanthosaurus would have extended for hundreds or thousands of square kilometers. The home range of a 5000 kg tyrannosaur might be proportionately higher. At some point, the size of the range needed to support the predator becomes too large for comfort. A territory of several thousand square kilometers cannot be effectively kept free of competitors, because patrolling and marking become impossible. When individuals are so far apart, locating a mate also becomes a major concern. Farlow & Holtz speculate that metabolic requirements for tyrannosaurs might be unexpectedly low, or prey productivity unexpectedly high. However, there are other solutions. Tyrannosaurs may have followed herds, for example, or taken advantage of some other behavior which ensured locally high prey densities and allowed boy to meet girl -- the Late Mesozoic equivalent of a freshman mixer.
... Well, perhaps not. But the actual provenance of tyrannosauroids remains at least a bit mysterious. Proposed closest relatives include Allosauridae, Acrocanthosaurus, Ornithomimosauria, and Therizinosauroidea. One of the best bets is Lambe's 1917) original intuition that tyrannosauroids were sisters of the Ornithomimosauria. Lambe should know, since he was the first to describe both Ornithomimus and Gorgosaurus. This is also the relationship most frequently recovered from cladograms. Holtz 2000) (reviewing previous work). Holtz ultimately concludes that ornithomimosaurs and tyrannosaurs probably form a monophyletic group, although his calculations do not yield a clean separation. In any case, that's the position we have taken here.
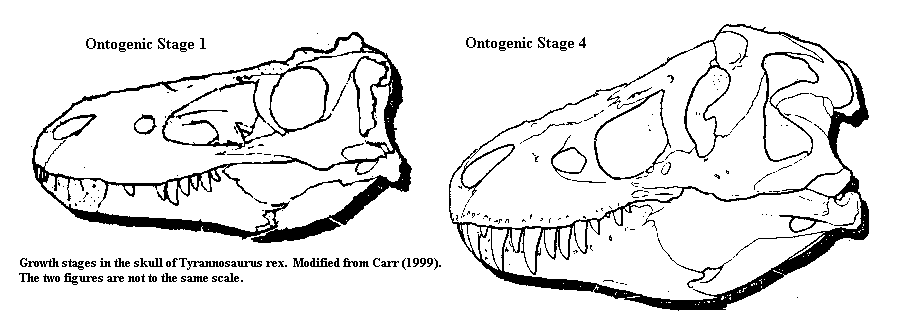
Much more recently, Carr (1999) has described a new, and disturbing, problem area in tyrannosaurid phylogenetics. It appears that many of the characters used to distinguish species and develop phylogenies are subject to considerable ontogenetic variation. That is, they change significantly over the life cycle of individuals. So, for example, a young Tyrannosaurus has a nasal with a smooth dorsal surface, while an old individual has a more rugose nasal. The first maxillary tooth changes from incisiform to conical, the postorbital shortens and develops a process running under the orbit, and the parietal develops a much larger nuchal crest. A review of the various characters, even synapomorphies, assigned to individual tyrannosaur species reveals that species differentiations are made on just such characters. So then, are we looking at artifacts of ontogeny or examples of paedomorphism and/or precociality? How do we score these age-related characters to calculate a cladogram? Some of the conceptual problems associated with ontogenic changes are discussed in connection with the work of Steyer 2000) on temnospondyl phylogeny. Carr's extensive list of character state changes suggests that this is a common and serious problem which needs to be addressed more carefully.
 Powers and Abilities Far Beyond Those of Mortal Men?
Powers and Abilities Far Beyond Those of Mortal Men?One of the relatively few undisputed ecological facts about the tyrannosauroids is that, whatever they did, they were the only ones who could do it. In most dinosaur-dominated faunas, a number of large predatory dinosaurs co-existed, and predator sizes graded down to successively smaller types. In the Late Cretaceous of North America and East Asia, the tyrannosauroids -- usually only one species at a time -- held the large predator job alone. The next largest carnivores were usually dromaeosaurs or other, much smaller theropods. Farlow & Holtz (2002). Thus, if tyrannosaurs were primarily scavengers, we must be willing to accept that these ecosystems had no large predators. That seems unlikely, to say the least.
One argument made by the proponents of scavenging is that the arms of tyrannosaurs were so useless as to preclude prey capture as a way of life. In response, Carpenter & Smith (2001) have built a good case that these diminutive arms were perhaps not so useless as they might appear. Carpenter & Smith map out the muscles on the pectoral girdle of Tyrannosaurus. While it is not necessary to get into the myological detail, C&S do effectively make the point that the girdles were large and well-muscled. The implication -- that the arms must have been useful for something -- is clearly brought home. Their discussion of the biomechanics of the arms is a little less convincing only because it is limited to the m. biceps. C&S attempt to work backward from the size of the tendon scar on the forearm to calculate the power and mechanical advantage of this muscle, the argument being that a strong biceps indicates an ability to hold struggling prey. The chain of reasoning -- all from the size of 70 million year old tendon marks on two individuals -- seems a bit attenuated. However, they mention two additional points which are better substantiated.
First, the arms are so constructed that adduction of the forearm (by the biceps) brings the hands toward each other. This tends to also to flex the digits, so that the most natural motion looks like clutching something close to the body.So far, so good. But, second, the range of motion is extremely limited. Its hard to say what this implies, but the restricted range of motion weighs strongly against the forearms being a prey-capturing device. It may also be worth noting that the area of origin for the m triceps is extensive, and the olecranon process of the ulna is notably large. Yet one of the specimens shows signs of an avulsion injury to the triceps origin. The triceps is a forearm extensor, the opposite of the biceps. Why should this muscle be particularly enlarged but, apparently, not yet large enough to withstand some severe strain placed on it? There are no obvious answers. We can only note that this is not what we might expect if the arms primarily served to immobilize prey.
So where does this leave the hunter-scavenger debate? Nowhere really. We have tentatively concluded that tyrannosaurs were slow. That conclusion is drawn from studies on flat ground, where tyrannosaurs and prospective tyranno-groceries were, so to speak, on a level playing field. But life, and terrain, are not so simple. One can only imagine tyrannosaur locomotion on broken slopes, terrain that would have given adult hadrosaurs (facultative quadrupeds, beautifuly designed for scampering uphill) an overwhelming advantage in mobility. Tyrannosaurs may have preferred juveniles and possessed dentition allowing them to make quick and highly efficient use of this resource. But juveniles are not always available. Large size and huge teeth always allow a carnivore to intimidate smaller killers, get a couple of quick bites in (ignoring bones), and amble off before the owners get organized. The arms could conceivably be used to subdue struggling prey, but don't they seem a whole lot better designed to contain and confine non-struggling, dead prey and keep it out of reach of smaller hunters while the tyrannosaur runs off with it? How about the range sizes? If tyrannosaurs needed a behavioral trick to concentrate food sources, how about following small groups of more efficient killers? The relationship need not have been purely parasitic. Four or five dromaeosaurs could probably harass and immobilize all but the largest herbivores of the Cretaceous, but even a mortally wounded hadrosaur would be unreasonably dangerous for the bird-boned predators. But, just about then, the local tyrannosaur would finally catch up and assume the minimal (to a tyrannosaur) risk of the final kill in exchange for first dibs on body parts. This kind of antagonistic commensalism requires no cooperation at all, but would naturally result in a predatory system of very high efficiency -- perhaps high enough to eliminate competition from traditional large predators. So, our provisional speculation is that tyrannosaurs were neither superpredators nor scavengers. They may have been more like thugs, living by a combination of smash-and-grab, extortion, gang violence and contract executions: not superpredators, and not scavengers -- but Sopranos. ATW030308
[1] I regret that I was unable to lay hands on a copy of Paul 2000). I understand that the argument for a swift, cursorial Tyrannosaurus is very well made in this piece. Interested readers should consult Paul, G 2000), Limb design, function and running performance in ostrich-mimics and tyrannosaurs. Gaia 15: 257-270.
[2] Could we fiddle with the angles to improve the results? Yes. However, the bottom line is that the angle is a function of speed. The faster Dino moves, the more bent over he is. The angles increase and the mechanical problem gets much worse. This, and some other questions are answered at Tyrannosaurus Was Not A Fast Runner
[3] Farlow and co-workers treat the head and body as rigid objects. They aren't. When the head and torso of the critter hit the ground, these portions decelerate very rapidly. But the tail and hindquarters are still moving at 20 m/sec. The combined torque, shear and compression effects are unpleasant to consider. They would almost certainly result in catastrophic spinal injuries. Tyrannosaurs are not well designed to perform somersaults.
Tyrannosauroidea:Definition: Tyrannosaurus > Tinamous. Includes: Itemirus, Labocania, Siamotyrannus, Stokesosaurus,
Range: Early Cretaceous to Late Cretaceous of North America, Europe, Mongolia, Central Asia & Southeast Asia.
Phylogeny: Coelurosauria : Compsognathidae + (Coelurus + Maniraptoriformes) + * : Eotyrannus + Tyrannosauridae.
Characters: skull large; jaw & neck muscle attachment areas expanded; prefrontal reduced or absent [S97]; upper temporal fenestrae separated only by sagittal crest [S97]; squamosal with pneumatic recess [H00*]; jaw articulation anterior to occipital condyle [H00]; palatines do not meet medially [H00]; palatal fenestra between palatine & pterygoid [S97]; premaxillary teeth U-shaped in cross-section [H00]; premaxillary teeth incisor-like with both carinae of each tooth being placed along the same plane perpendicular to the main axis of the skull [H00*]; semilunate carpal block reduced or absent [H00*]; ischial foot absent [S97]; semicircular scar on posterior face of ischium {H00*]; long tibia & metatarsi; primitively, medium-sized predators.
Notes: [1] according to Holtz, Stokesasaurus and Itemirus are a separate clade, which shares very deep pockets on the lateral surface of the basipterygoid process. [2] the characters ascribed to [S97] are those which unite tyrannosaurs and Maniraptora. The characters ascribed to [H00*] are those which unite tyrannosaurs and ornithomimosaurs.
Links: Tyrannosauroidea Tree of Life); Tyrannosauroidea Mikko); tyrannosauroidea.htm; GEOL 104 Lecture 23- Theropoda II- Tyrants, ostrich mimics, and ...; Thomas R. Holtz, Jr..
References: Holtz (2000) [H00], Sereno (1997) [S97]. ATW021210.
 Eotyrannus: E. lengi Hutt et al. 2001.
Eotyrannus: E. lengi Hutt et al. 2001.
Range: Early Cretaceous (?Barremian) of Europe (England).
Phylogeny: Tyrannosauroidea: Tyrannosauridae + *.
Characters: ~5m (subadult); premaxilla small [H+01]; premaxilla anterior border almost vertical [H+01]; premaxilla taller than long [H+01]; maxillae with anterior portion laterally flattened [H+01]; maxilla having sharp lateral border with antorbital fossa [H+01]; maxilla ventral edge straight [H+01]; nasals fused [H+01]; nasals dorsally concave [H+01] [1]; quadrate robust [H+01]; dentary thin, internally flat, with parallel dorsal & ventral borders having a slightly convex outline in lateral view [H+01]; teeth with denticulations continuing across the crown [H+01]; premaxillary tooth row oriented more or less transversely [H+01]; premaxillary teeth smaller than lateral teeth & D-shaped in cross-section [H+01]; premaxillary teeth with serrated carinae [H+01]; premaxillary teeth with straight, conical roots [H+01]; maxillary teeth carinate, with anterior denticles smaller than posterior [H+01]; lateral teeth not laterally expanded [H+01]; neural spines low, transversely thin & extend entire length of centrum [H+01]; postzygapophyses directed posteriorly & flaring laterally to twice width of prezygapophyses [H+01]; anterior dorsal vertebrae with elongate centra, constricted at midlength and flared at ends [H+01]; dorsal centra shorten towards sacrum {H+01]; sacrals with pleurocoels; scapula strap-like, with distal expansion (if present) not preserved [H+01]; acromion and glenoid much wider than scapular shaft [H+01]; coracoid with circular glenoid facing posteriorly (??) [H+01]; boss-like coracoid tubercle 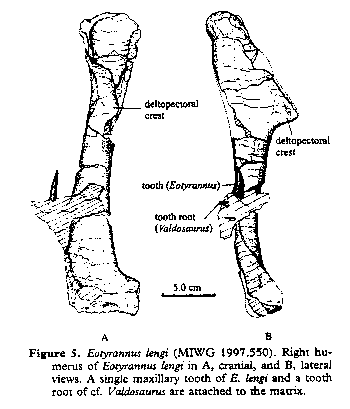 present on ventral part of lateral surface [H+01]; arms fairly long; humerus 2/3ds length of scapula (23 cm) [H+01]; humeral head well-separated from shaft [H+01]; humerus with very large deltopectoral crest terminated abruptly at ~40% shaft length [H+01]; humerus with large internal cavity dorsally & smaller cavities ventrally just distal to deltopectoral crest [H+01]; carpals not reduced [H+01]; tridactyl hand [H+01]; Mc I very long (other Mcs incomplete) [H+01]; hands elongate (manus II is 95% of humerus length) and slender [H+01]; unguals also elongate, curved and laterally compressed [H+01]; hindlimbs gracile {H+01]; cnemial crest poorly developed [H+01] fibula with proximal crest & large foramen just distal to crest [H+01]; Mt II slender with expanded proximal head & medial facet for Mt III; Mt IV similar, with convex dorsal surface
present on ventral part of lateral surface [H+01]; arms fairly long; humerus 2/3ds length of scapula (23 cm) [H+01]; humeral head well-separated from shaft [H+01]; humerus with very large deltopectoral crest terminated abruptly at ~40% shaft length [H+01]; humerus with large internal cavity dorsally & smaller cavities ventrally just distal to deltopectoral crest [H+01]; carpals not reduced [H+01]; tridactyl hand [H+01]; Mc I very long (other Mcs incomplete) [H+01]; hands elongate (manus II is 95% of humerus length) and slender [H+01]; unguals also elongate, curved and laterally compressed [H+01]; hindlimbs gracile {H+01]; cnemial crest poorly developed [H+01] fibula with proximal crest & large foramen just distal to crest [H+01]; Mt II slender with expanded proximal head & medial facet for Mt III; Mt IV similar, with convex dorsal surface
Notes: [1] it appears that the medial branch of the usual three-pronged posterior end of the nasal is longer than the lateral branches.
Image: Right: from Site-No-Di, by Mituse Komiyagi (I think). Left: from [H+01].
Links: DinoData Dinosaurs E079 EOTYRANNUS lengi; Eotyrannus Fact Sheet - EnchantedLearning.com; DinoWight- Eotyrannus, an Isle of Wight Tyrannosaurid; Eotyrannus lengi Dutch); Eotyrannus attacking Hypsilophodon; Eotyrannus Dutch); ???????-eotyrannus Japanese); USATODAY.com - Ancestor of T-rex discovered in Britain; Isle of Wight & Natural History Museum Dinosaur Displays.
References: Hutt et al. 2001) [H+01]. ATW021210.
checked ATW050619