| Coelurosauria | ||
| The Vertebrates | Tyrannosauridae |
| Vertebrates Home | Vertebrate | Vertebrate |
|
Abbreviated Dendrogram
Dinosauria
├─Ornithischia
└─┬─Sauropodomorpha
└─Theropoda
│
└─Coelurosauria
├─Tyrannosauroidea
│ ├─Eotyrannus
│ └─Tyrannosauridae
│ ├─Alectrosaurus
│ └─Tyrannosaurinae
└─Maniraptora
├─Therizinosauroidea
└─┬─Alvarezsauridae
└─┬─Oviraptorosauria
└─┬─┬─Troodontidae
│ └─Dromaeosauridae
└─Aves
|
Contents
Index |
Considering the various lines of theropods, it is interesting that the most primitive forms have very long tails, long bodies, relatively small heads, short hind legs, and relatively large forelimbs. The later and more advanced types had relatively shorter bodies and tails, larger heads, larger hind legs, and (in the case of the giant forms) smaller and smaller forelimbs. This evolutionary trend reaches its culmination in the Tyrannosauridae of the late Cretaceous of the Asiamerican landmass.
At this time a narrow north-south seaway divided what is now North America into two large islands; the same thing happened in Asia. But a land-bridge linked east Asia (Mongolia, China, etc) with western North America. Apart from one very primitive form from India (then an island), it was only on this Cretaceous continent, called "Asiamerica", that the more derived tyrannosaurs are found .
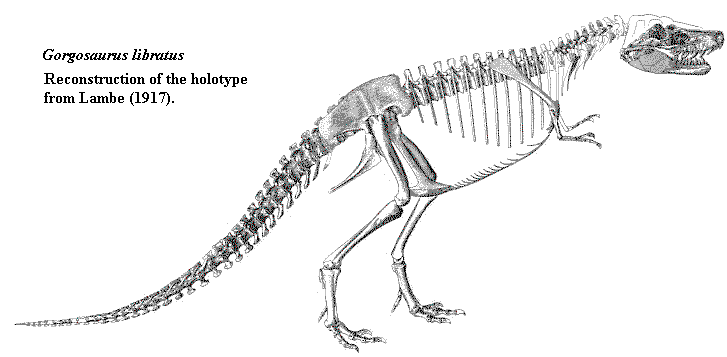
Although old drawings of Tyrannosaurus rex give the beast an almost erect stance and a long reptilian tail, which dragged on the ground. This is now thought to be incorrect. Lambe's 1917) illustration at right was actually somewhat ahead of its time. In fact, in his description of Gorgosaurus he speculates that the animal dragged its tail little, if at all. It is now believed that back was held horizontally, as in other theropods (although the animal could likely rear up when the situation required); and the tyrannosaur tail was, as a proportion of overall length, the shortest of any large theropod. The fearsome eagle-taloned feet of the tyrannosaurs are also a myth; the claws on the hind feet were short, rounded, and hoof-like. All the killing was done by the huge jaws with their array of 15-centimetre steak-knife- like teeth. The power of these jaws has been the subject of a great deal of research, and was almost incredible. Think of a huge terrestrial Great White shark. The animal could bite out a wound a metre long and well over thirty centimeters deep and wide. Yet only with such ferocity could it possibly take on prey like the elephant-sized horned dinosaur Triceratops -- if it did. Jack Horner (and very few others) has strongly argues that the big tyrannosaurs were almost exclusively scavengers, using their obvious power to intimidate smaller killers, rather than hunting themselves. Recent work by John Hutchinson and others have suggested that Tyrannosaurus was a relatively slow animal. If correct, this would be consistent with a role as a Mesozoic extortionist, rather than a hunter.
One thing that is quite bizarre about the tyrannosaurs are their tiny little forelegs [1]. Despite their diminutive status, they clearly served some purpose, indicated by the the relative robustness of the arms and repeated stress fractures [2]. But generally, like ground-living large running birds, the big theropods would have acted more like huge flightless birds of prey than the overgrown bipedal lizards of popular imagination. Interestingly, after the extinction of the dinosaurs, a number of real flightless birds - Diatryma, Phorusrhacos, etc - took over the ecological niche of the theropods, becoming huge-headed, fast-running predators, 1.5 to 2 metres or more in height.
Although the derived tyrannosaurids have sometimes been divided into two subfamilies: the small primitive aublysodontines and the large tyrannosaurines, but the fragmentary material of the former means they might not constitute a valid taxon. In fact Aubylson is believed to be a misinterpretation of juvenile tyrannosaurine material. All are known only from fragmentary remains. They are based almost entirely on distinctive nose and ridged premaxilla teeth (in the front of the jaw). It has been plausibly argued that the teeth are just typical teeth of young tyrannosaurids. Further, whilst Aublysodon-like teeth are fairly common and found in practically all horizons where tyrannosaurid body fossils are found, not one adult theropod skull is known from North America with such a tooth actually in place in a premaxilla. The best-known aubylsodont, the so-called "Jordan theropod" (Aublysodon molnari [Paul 1988)], = Stygivenator molnari [Olshevsky (1995)]), actually just a snout with some teeth, has been variously interpreted as Aublysodontine (Paul, Holtz), dromaeosaurid, and tyrannosaurid, but is most likely a juvenile (length about 3.8 meters, weight around 200 kg) specimen of T. rex. MAK990512 revised 121205
When is Tyrannosauridae not Tyrannosauridae? Answer: when its Deinodontidae
Earlier, mention was made of Podokesauridae, the more correct senior synonym of Coelophysidae/Coelophysoidea.
Now it seems that the more correct term for Tyrannosauridae is Deinodontidae. To quote Matthew Martyniuk
Deinodontidae (Cope 1866)...was in clear, widespread use through the mid 20th Century (as in Maleev 1955) and almost always treated as the senior synonym of Tyrannosauridae (Osborn 1905). However, Russel (1970) argued that Deinodontidae be abandoned, because he considered the type specimens of Deinodon (isolated teeth) not diagnostic, rendering the name a nomen dubium. However, the teeth are clearly diagnostic at the family level and possibly even genus and species, as they must have come from either Daspletosaurus or (more likely) Gorgosaurus, and the rocks those dinosaurs come from are well enough sampled to rule out the presence of a third large tyrannosaur species unless such compelling evidence is found. Similarly, it is questionable whether or not the pertinant ICZN rules allow for abandoning a name due to a dubious type genus. Even if this is the case, it is only followed sporadically in the literature, and many family names remain in use that are based on dubious type material, including Hadrosauridae, Ceratopsidae, and Troodontidae (the latter is also based exclusively on teeth of questionable diagnosability at the genus and species levels). Olshevsky (1991) recognized this, but argued that the name is still invalid because Cope initially spelled it Dinodontidae, and the name Deinodontidae was an emended spelling not published until 1914, after Tyrannosauridae. He concluded that therefore Deinodontidae (with an e) is a junior synonym and Dinodontidae (no e) is a nomen oblitum. However, Olshevsky's argument is incorrect because the ICZN clearly mandates that any family names based on misspellings or unjustified spelling changes of their type genus (Cope spelled the name Dinodon) can and must be emended by any subsequent revisor, and that this does not change the original authorship or date of the name (ICZN Article 35.4.1). Also, note that even if Deinodontidae and Deinodontoidea are ignored, several studies have found Coelurus fragilis to be a "tyrannosauroid", and so the next available name for that group after Deinodontoidea is Coeluroidea (Marsh 1881).
Were we to be totally consistent and rigorous, we would have to stop using Tyrannosauridae and replace it instead with Deinodontidae, (and Deinodontinae and Deinodontoidea. Or even, for that last, Coeluroidea). The present author for one (and I know I am not alone here) has never gotten over the loss of the marvellously imaginative Brontosaurus ("thunder lizard") in favour of that insipid Apatosaurus ("deceptive lizard"). And while "terrible tooth" is a cool name, it is still no substitute for "tyrant lizard". For this reason -, and while acknowledging it is not technically correct - we have opted to retain Tyrannosaur-inae/idae/oidea as the names for those Linnaean and Cladistic taxa, of which the great king of the tyrant lizards is both the nomenclatural and evolutionary culmination. - MAK120209
Notes
[1]The present author (MAK) had originally suggested that tyrannosaur forelimbs became smaller as the creatures evolved, and would have dissappeared entirely had their evolution not been cut short by the terminal Mesozoic catastrophe. Two inspirations for this claim were Greg Paul (1988) and Dougal Dixon (1988). However, a correspondent points out in an email (121202) that this hypothesis is not supported by more recent research. As he explains, "Of the genera ascribed to the family tyrannosauridae, Currie points out that the different genera have forelimbs of the same size (the exception of the rule is Tarbosaurus, which have forelimbs smaller than the "standard" size), (Currie 2003) although there can be some differences in hand bone proportions in different genera, as pointed out by Holtz 2004. One should also remember the morphological variation seen in animals of closely related, and even within the same genera....Gregory Paul also seems to have changed his view on the subject. He have told me (email November 2012) that larger sample size currently seems to speak for a forelimb size stable over geological time (rather than the idea of gradual forelimb reduction) and believe there are new unpublished data which might could settle the issue. "
[2] More on forelimb function comes from Lipkin and Carpenter 2008 who compared the forelimbs of different tyrannosaurids (Gorgosaurus, Albertosaurus, Daspletosaurus and Tyrannosaurus).
Tyrannosauridae: Definition: Tyrannosaurus + Aublysodon. Includes: Alioramus, Aublysodon, Daspletosaurus, Tarbosaurus, Tyrannosaurus.
Range: Early Cretaceous to Late Cretaceous of North America, Europe, Mongolia, North China and Japan [M99]
Phylogeny: Tyrannosauroidea: Eotyrannus + * : Alectrosaurus + Tyrannosaurinae.
Characters: Large head [C97]; skull akinetic, laterally compressed, boxy in lateral view [C97]; curved jaws (brings all teeth to bear at same time, as opposed to slicing -- compare Prosauropoda or Coelophysis); mandibular symphysis cartilaginous; ventral ramus of premaxilla taller dorsoventrally than long rostrocaudally [$H01a]; marked ventral curvature of maxilla [$H01a]; snout deep [C97]; nasals fused [C00] [$ H01a]; nasals narrow caudally behind external nares; orbits large [C97]; possibly stereoscopic vision, at least in some [C97]; snout surface rugose [C00]; rugose dorsal, preorbital surface of lacrimals with bone "horn" in some species; prefrontals reduced [$?H01a]; lacrimal-frontal contact present; supratemporal fossa occupies at least the posterolateral third of frontal [$H01a]; well-developed sagittal crest on dorsal surface of parietals [$H01a]; lateral nuchal crest formed by parietals [$?H01a], jugal & quadratojugal deeply invade lateral temporal fenestra [CC00]; at least twice as tall as foramen magnum vertical height [C00]; quadrate height 40-50% of skull height at mid-orbit; quadrate and/or articular invaded by air sacs connected to middle ear [W97]; squamosal- quadratojugal flange intruding anteriorly into infratemporal fenestra, nearly contacting rostral margin [$H01a] [$MB89]; squamosal with large pneumatic space [W97]; pair of tab-like processes on supraoccipital wedge [$?H01a]; optic & olfactory nerve tracts enlarged [C97]; rostral portion of fused vomers expanded to greater than twice shaft width; in some, palatine dorsal surface without prominent muscular fossae [$?H01a]; air sinus in ectopterygoids is complex & reticulated [W97]; basisphenoidal sphenoidal sinus deep, with large foramina [$?H01a]; supradentary fused to coronoid, restricting lateral motion of intramandibular joint; caudal surangular foramen very large [$H01a] [$MB89]; retroarticular process of articular absent [$H01a]; premaxillary tooth row arcade more mediolaterally than rostrocaudally oriented [$?H01a]; premaxillary teeth incisiform, D- or U-shaped in cross-section with median ridge on caudal surface [C00] (or with two carinae "in same plane perpendicular to the skull axis" [$?H01a]); dentition heterodont; premaxillary teeth smaller than lateral teeth [$H01a]; incisiform premaxillary teeth; large, curved, serrated teeth; lateral teeth incrassate and capable of crushing bite [C00]; probable large wattle on neck [C97]; short, heavily-built neck [C97]; cervicals short & wide (shorter, stronger neck related to large head) [C97]; cervical air sacs pneumatize vertebrae at least as far anteriorly as 2nd cervical [W97]; neural spines short & robust (muscle & ligament attachments to support large head) [C97]; presacral centra axially short [C00]; body deep & short [C97]; gastralia from coracoids to pubis [C97]; distal caudal neural spines axially elongate (what does this mean?) [$?H01a]; forelimb reduced [$MB89]; scapula contributes more than half of the glenoid [$?H01a]; acromion expansion well-developed, more than twice scapula midshaft width [$?H01a]; scapular-humerus ratio between 2.2 and 2.5 [$?H01a]; humerus greatly reduced; femur-humerus ratio between 2.8 and 3.5 [$H01a]; radius, distal end flattened anteroposteriorly; fibular cranial tubercle distal to cranial expansion composed of two longitudinal ridges [$H01a]; distal carpals of adults poorly formed and lack trochlear surfaces [$?H01a]; metacarpal III very reduced and bears no digit [$?H01a]; manus with 2 digits [$MB89]; manus I curves away from II [C97]; pollex (I) ungual roughly equal to ungual of digit II in size; horizontal medial shelf from preacetabular blade to sacral ribs [$?H01a]; preacetabular notch on ilium [$?H01a]; broad ventral hook-like projection from preacetabular blade of ilium [$?H01a]; supraacetabular crest on ilium reduced [$?H01a]; dorsal surfaces of iliac blades very close to the midline [$?H01a]; ilium long, slight shorter than femur [$?H01a]; distal end of iliac pubic peduncle oriented horizontally; pubes with large "boot", about half as long as pubic shaft [$?H01a]; ischium shaft long but more slender and shorter than pubis [$?H01a]; pronounced semicircular scar on the anterolateral surface of the ischium, just distal to iliac process [$?H01a]; ischial foot absent (pointed tip) [$?H01a] [$MB89]; femoral anterior trochanter as tall as greater trochanter; lesser trochanter at least as tall as femoral head [$?H01a] probably same as previous character); anterior and greater trochanter junction at distal end of anterior trochanter; long ascending process of astragalus [C97]; tibiae and metatarsi elongate relative to other theropods of same femoral length [$?H01a]; fibula with large, bipartite protuberance on anterior face, near proximal end [$MB89]; presence of an arctometatarsus (proximal end of metatarsal III reduced to a solid splint) [C00] [$?H01a]; dorsal surface of metatarsal III crescentic and restricted to distal portion of the metatarsal dorsal surface [$?H01a]; metatarsals II and IV contact at mid-shaft on plantar surface; 3 substantial forward pointing pedal digits with large claws; small round or hexagonal scales on tail.
Notes: [1] tooth fractures very common in this group, with distal crowns snapped off sometimes in the majority of teeth. The root was apparently retained in the mouth for considerable periods of time. This may be related to bone-crushing as a common practice [B97]. Particularly high degree of cranial bone pneumaticity may also be associated with crushing, as this allowed strong, but light bones having a considerable degree of flex in response to mechanical stress. [W97]. This differs from the construction of, for example, carnosaurs, which tended to solid bones but with considerable cranial kinesis. Thus, in carnosaurs, flexibility was achieved by permitting the bones to move relative to each other. In tyrannosaurids, the bones were fixed in position, but were designed to bend. [C97].
Links: Welcome to the UCMP T. rex Expo!; The Story and Contest of Dinosaur Stan the Tyrannosaurus rex; Discovery of a furcula ...; Tyranosaurids; Review of the Tyrannosauridae; Tyrannosauridae; Tyrannosaurids; What's the difference between Tyrannosauridae, Tyrannosaurinae, etc.?; Tyrannosauroidea; Tyrannosaurids.
References: Buchholtz 1997) [B97]; Carpenter (1997c) [C97]; Currie (2000) [C00]; Currie & Carpenter (2000) [CC00]; Holtz (2001), Mader & Bradley (1989) [MB89], Manabe (1999) [M99], Witmer (1997) [W97]. ATW060214.
 Alectrosaurus: A. olseni Gilmore 1933.
Alectrosaurus: A. olseni Gilmore 1933.
Introduction: Alectrosaurus is a stand-in here for a poorly known clade frequently referred to as Aublysodontinae. Alectrosaurus itself was founded on a partial leg and some forelimb material, leftover gleanings from the Third Mongolian Expedition. The long arms are now believed to be unrelated portions of a therizinosaur [MB89]. However, these same forelimbs look a bit like the arms of Eotyrannus [H+01]. Perhaps this issue is worth a second look. There is not much phylogenetic distance between a basal tyrannosaurid and a basal therizinosaur. Perle referred additional material to this taxon in 1977. This reference has been accepted by some [e.g. H01a], but questioned by others [MB89].
Range: Late Cretaceous of China, Mongolia & Central Asia?
Phylogeny: Tyrannosauridae: Tyrannosaurinae + *.
Characters: ~5m? 8-9m?; long, low, sloping head [C00] [H01a]; nasals smooth [C00]; $ premaxillary teeth lacking serrations [C00] [H01a]; $ premaxillary teeth with prominent vertical ridges on lingual surface [H01a]; first 2-3 maxillary teeth incisiform [$H01a] [C00]; 17 maxillary & 19 dentary teeth (more than Tyrannosaurines) [C00] [H01a]; lateral teeth narrower & more blade-like than in Tyrannosaurinae [C00]; femur & tibia of about equal length [MB89]; femur, medial distal condyle with spike-like dorsal process [CT05]; tibia with stout, laterally curved cnemial crest [MB89]; distal tibia expanded laterally & flattened anteroposteriorly [MB89]; distal tibia with ridge articulating with groove on astragalus (as in all Tyrannosauridae) [CT05]; fibula with proximal end much expanded anteroposteriorly, with large concavity on medial surface [MB89]; fibula very thin below proximal expansion [MB89]; fibula anterior surface below proximal expansion with protuberance formed from 2 lateral ridges [MB89]; astragalus with strong ventrolateral buttress just proximal to trochlea [MB89] [CT05]; calcaneum taller than wide, narrow in anterior view [MB89] [CT05]; distal articular surfaces of metatarsals II & IV, and of most phalanges, elevated on pedicles [CT05]; Mt I much reduced (13% of Mt IV length) [MB89]; Mt II & IV unusually thin for length [MB89]; MtII with seep fossa for medial collateral ligament [MB89]; Mt III strongly compressed proximally (arctometatarsus) [MB89]; distal condyle of Mt III surrounded by a marked lip [MB89]; Mt V vestigial, laterally compressed & curves anteriorly at distal end [MB89]; phalangeal formula 23450 [MB89]; phalangeal collateral ligament fossae well-developed [MB89]; all phalangeal articulations hinge-like [MB89]; flexor tubules on pedal phalanges large and posteriorly extensive [MB89] pes III shorter relative to Mt III than in other tyrannosaurids [MB89].
Image: from Alectrosaurus.
Links: Alectrosaurus olseni; Paleontology and Geology Glossary; Alectrosaurus Korean); Nanotyrannus; Gigadispute; Alectrosaurus Specimens.
References: Carr et al. 2005) [CT05]; Currie (2000) [C00]; Holtz (2001a) [H01a]; Hutt et al. (2001) [H+01]; Mader & Bradley (1989) [MB89]. ATW031029, revised ATW080117.
checked ATW080118, last modified MAK121205