Synapsida
Caseasauria
Taxa on This Page
- Caseasauria X
- Caseidae X
- Eothyrididae X
The Eothyrids
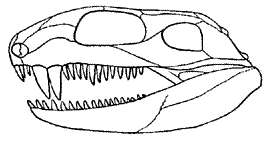
This family of very primitive pelycosaurs includes two species, known only from several skulls and limb bones from the Early Permian of North America. Eothyris, from the Early Permian of Texas (Belle Plains formation, Artinskian age), is known from a single skull (right), 62mm in length. The teeth are simple conical structures, and there are distinct caniniform teeth. Oedaleops, from the Early Permian of New Mexico (Abo Cutler Formation, Sakmarian , is known from three partial skulls and some limb elements.
In terms of cranial morphology the eothyridids are the most primitive of any pelycosaur. For this reason they are, in modern cladistic classifications, considered a sort of ancestral group (or rather, the sister group to all other synapsids). However they appear quite late in the fossil record (they are one of the last groups of pelycosaurs to appear). This means either that they existed for a long time but in upland areas or isolated geographic regions away from any chance of fossilisation, or, alternatively, that they are only secondarily primitive. Either hypothesis is valid, and the answer, if it is ever known, must await further fossil discoveries.
The eothyririds share a few unique characteristics (synapomorphies) with the caseids, indicating they may be ancestral to the latter. The two groups are united in the clade (suborder?) Caseasauria. (MAK
000720).
-- and the Synapsid Skull
Sufficiently far back, all of us can trace our family tree back to some bad-tempered sail-back, Dimetrodon-like sphenacodont pelycosaur. Except for an annoying mess in the Late Jurassic to Early Cretaceous eras, the pedigree is fairly clear all the way from the Early Permian to my great-great grandfather, whose explanations of his ancestry and reason for being in Shanghai were (shall we say) a little unclear. However, aside from Mesozoic muddles and mendacious mercenaries, it is a remarkably straight line.
Moving beyond the Early Permian is much more difficult. A number of pelycosaurs are known from the Late Carboniferous, but they all seem rather specialized. A number of other scrappy specimens have turned out not to be pelycosaurs at all. The most morphologically basal synapsids we know reasonably well are the eothyrids from the Lower Permian of North America. The eothyrids consist of two adequately known specimens, Eothyris Romer and Oedaleops Langston.
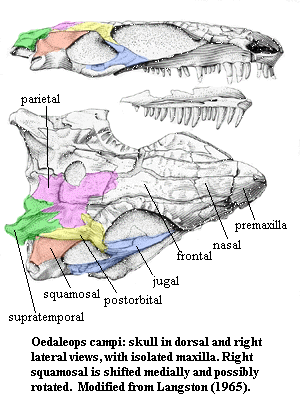 The Eothyrids look more or less like Bob the Basal Amniote, whose osteological adventures are sometimes chronicled in the Bones section of this site. That is, most of the skull is composed largely of a series of long dermal bones, rather like the hull of a well-built sailboat. Pressure at any point is flexibly distributed over a wide area while the overall structure remains rigid. The eothyrids have none of the mammalian innovations. The skull is flat -- even depressed according to Langston (1965). The jaw articulation is at about the same level as the tooth row. In fact, there is almost nothing to suggest its distant mammalian affinities except perhaps the curvature of the lower jaw and the shape of the rostrum.
The Eothyrids look more or less like Bob the Basal Amniote, whose osteological adventures are sometimes chronicled in the Bones section of this site. That is, most of the skull is composed largely of a series of long dermal bones, rather like the hull of a well-built sailboat. Pressure at any point is flexibly distributed over a wide area while the overall structure remains rigid. The eothyrids have none of the mammalian innovations. The skull is flat -- even depressed according to Langston (1965). The jaw articulation is at about the same level as the tooth row. In fact, there is almost nothing to suggest its distant mammalian affinities except perhaps the curvature of the lower jaw and the shape of the rostrum.
What is somewhat different about the skull is, of course, that it is a synapsid skull. That is, it has only one temporal opening on each side. The opening is defined very simply by the jugal, postorbital and squamosal -- at least it probably is. The squamosal is displaced or damaged on both sides of the skull and both posterolateral quadrants of the skull are somewhat distorted, as can be seen in the figure. (For this reason the quadratojugal appears to participate in the margin of the temporal fenestra.)
In diapsids, a second, dorsal fenestra opens at the junction of the squamosal, parietal and post orbital. No such junction exists in the eothyrids because the squamosal does not extend onto the top of the skull. Instead, an enlarged supratemporal intervenes and forms a sharp projection at the corner of the skull table. As if to compensate -- and perhaps it did -- both the orbits and the pineal foramen are greatly enlarged.
It would not be safe to assume that the synapsid temporal fenestra is strictly homologous to the lower temporal fenestra of diapsids. Although they occur at the same intersection, a number of lineages radiated from the ancestral amniote stock with different patterns of fenestration (see, e.g. Procolophonia), and it is entirely possible that the synapsid and diapsid lineages are separately derived from within Amniota. In fact, that is the usual assumption. If anything, the synapsids seem more closely related to anapsids than diapsids. Upper Carboniferous amniote fossils are few and far between, so that the details of the amniote radiation have remained elusive.
If the diapsids were so successful with their comparatively light-weight, strongly fenestrated skulls, how were the Synapsids able to compete? After all, the earliest synapsids had not yet developed the jaw specializations which enabled sphenacodonts and more derived forms to make good use of the comparative solidity of the synapsid posterior skull. Thus, they had only half the tooth row and perhaps half the jaw muscle strength of a well-developed diapsid.
Here we enter a more speculative area. Perhaps the answer relates to the odd flange of the frontal that enters the margin of the orbit at the top of its arc, together with that other singular feature, the shortened, highly arched ("inflated") rostrum. In a somewhat different form, we have seen this "bulldog syndrome" before in odd creatures like the porolepiforms, guildayichthyiforms, sphenodonts, and perhaps in some of the anthracosaurs and cichlids. In each case, we find some combination of the following features: the tooth row is fairly short, the orbits are large, anterior, and closely spaced, caniniform teeth are found on the anterior tooth row, and the medial bones are strong, uncomplicated and sometimes fused medially. As Lund (2000) points out in the extreme case of guildayichthyiform fish, this syndrome is associated with skulls which are arched and transfer stress from the anterior jaws medially to the rear of the skull
-- literally like bulldogs, which sacrifice speed and big jaws for incredible strength in the anterior jaw.
Perhaps what Langston sees as skull depression is actually the osteological remains of a peculiar skull kinesis which allowed the median arch of the skull to be laterally reinforced even as it flexed slightly and locked on to some food source too tough or too uncooperative for normal diapsid slash and gulp tactics. The lateral flanges of the frontals and parietals are then easily explained as stiff crossbars to keep the skull from collapsing inward too far. For these tactics, an eothyrid would need an upper temporal fenestra like -- well, like a hole in the head. ATW 010601
Caseidae

The caseids were a widespread and diverse group of herbivorous pelycosaurs, which appeared during the later Early Permian and persisted until the late Middle Permian. Although ranging in size from 1 to 6 meters in body length, caseids are surprisingly conservative in their skeletal anatomy and body proportions. All were massively built animals with small heads and barrel-like bodies. Animals like Angelosaurus and Cotylorhynchus, which exceeded 4 meters in length and were the largest of the pelycosaurs, seems to have played the same ecological role during the later early Permian that the pareiasaurs and herbivorous dinocephalians did during the Middle and Late Permian. Caseids and eothyridids share a number of specialised features associated with the morphology of the snout and external naris and it is likely that the latter evolved from the former. The two together form the clade Caseasauria.
The caseid skull is distinctive in the presence of relatively large temporal openings, enormous external nares (nostril openings, probably to house some kind of sensory or moisture-conserving organ), and large pineal opening, and a snout or upper jaw that dramatically overhangs the tooth row to form a forward projecting rostrum. The external surface of the skull is skulpted with rounded deep pits and sometimes crevice-like depressions. The marginal teeth are quite similiar to the teeth of parelasaurs.
In contrast to most other pelycosaurs, except Edaphosaurus, and as befits the herbivorous lifestyle, the teeth are remarkably uniform. There is a general reduction in the number of marginal teeth cheek teeth. All caseids, whether modest in size, or enormous, are characterized by small cervical vertebrae, bulky, barrel-shaped bodies and relatively massive limbs. There is reduction in the phalangeal formula, ranging from 2-3-4-4-3 to 2-2-2-3-2. In Corylorhynchus the claw bearing element is very large and broad, with apparently sharp edges on the sides. While in Angelosaurus the claw bearing phalanges are also massive, but more conservative in morphology, roughly triangular in outline. This may indicate adaptations to different types of terrain. MAK.
Descriptions
Caseasauria: Nitosaurus?
lwP-upP of NAm, Eur & Russia.
Dentary with long, ascending untoothed posterior process; $ long external nares with external narial shelf ; short preorbital skull; pointed rostrum formed by dorsal process of premaxilla; frontal contribution to orbit small (primitive) through narrow lateral process; broad, low skull table; large pineal foramen; wide supratemporal (primitive); slanted occipital plate.
Links: Synapsid classification and autapomorphies; Biology 356; Introduction to the Pelycosaurs.
References: Langston (1965). 010531.
 Eothyrididae: Eothyris, Oedaleops.
Eothyrididae: Eothyris, Oedaleops.
Small to medium (<1m?), primitive pelycosaurs, possibly polyphyletic as including caseids. Short antorbital region; teeth simple cones except $ large caniniform teeth on premaxilla and near anterior of maxilla; groove and/or foramen above canines; tooth row horizontal and on same level as jaw articulation; ventral surface of lower jaw rounded; no retroarticular process; parasphenoid ventral plate broad; quadrate ramus of pterygoid has ventral shelf; pointed muzzle formed by a dorsal process of the premaxilla; rostrum formed by premaxilla & nasal overhangs jaw; nares large; maxilla with narial (not nasal) flange; lacrimal reaches nares anteriorly and jugal posteriorly; quadratojugal visible in lateral view; quadratojugal meets maxilla and excludes jugal from ventral margin of skull; postorbital large and extends far posteriorly; cranium generally broad and perhaps depressed (not domed); frontal has lateral process participating in orbit; parietal broad, with very large pineal foramen; parietal forms lappets into postorbital; broad supratemporal; supratemporal "horn"; postparietal not fully fused (??); broad supraoccipital; occiput formed from large, flat supraoccipital fused to paraoccipital; post-cranium unknown except limb fragments; probably insectivorous and/or carnivorous.
Links: PELYCOSAUR
- Paleontology and Geology Glossary; Synapsid classification and autapomorphies; Biology 356; Schnellbestimmung anhand von typischen Schädelmerkmalen (German); reptiles; Oedaleops. .
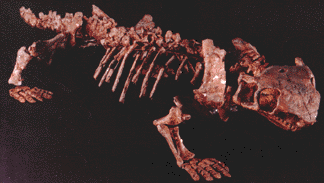 Caseidae: Angelosaurus, Casea, Caseoides, Caseopsis, Cotylorhynchus, Ennatosaurus, Phreatophasma.
Caseidae: Angelosaurus, Casea, Caseoides, Caseopsis, Cotylorhynchus, Ennatosaurus, Phreatophasma.
Early Permian to Late Permian of North America, Europe & Russia.
1 to 4m herbivorous pelycosaurs with "slanted" skulls. Skull small; skull ornamented dorsally with small pits and some crevices; palatal denticles reduced; denticles (not teeth) absent on lower jaw; $ marginal teeth generally foliate (bearing multiple cusps); no caniniform teeth; largest teeth anterior on both jaws; snout (premaxilla) strongly arched upward and overhangs tooth row to form anteriorly projecting rostrum; $ large, high nares with external narial shelf; large pineal foramen; skull table broad & low; temporal opening very large; dorsal spines absent; small cervical vertebrae; very large rib cage and robust scapula; 3-4 sacral ribs; all limbs robust; phalangeal count reduced, as is size of feet; apparently successors to edaphosaurids as most common herbivorous pelycosaurs.
Image: Ennatosaurus from mathematical.com by permission.
Links: Introduction to the Pelycosaurs; PELYCOSAUR
- Paleontology and Geology Glossary; Synapsid classification and autapomorphies; dinosaurs-ennatosaurus; Biology 356 (outstanding site!); Synapsida; Stars of the Show (A to I); Schnellbestimmung anhand von typischen Schädelmerkmalen (German); Caseidae Mikko's Phylogeny); Stars of the Show (A to I) Ennatosaurus skull).
References: Langston (1965). ATW030614.