Lanarkia: a thelodont, showing the flattened, unarmoured body typical of this group.
Graphic from Faktaside om fossile urfisk fra Norge, © 1998 Paleontologisk museum - University of Oslo.
| Thelodonti | ||
| The Vertebrates | Thelodonti |
| Vertebrates Home | Vertebrate | Vertebrate |
|
Abbreviated Dendrogram
Vertebrata
├─Conodonta
└─┬─Pteraspidomorphi
└─Thelodonti
├─┬─┬─Nikoliviidae
│ │ └─Furcacaudiformes
│ └─Thelodontida
└─┬─┬─Katoporida
│ └─Cephalaspidomorphi
│ ├─Galeaspida
│ └─┬─Pituriaspida
│ ├─Osteostraci
│ └─Gnathostomata (conventional placement)
└─┬─Loganiidae
└─Gnathostomata (alternate placement)
├─Placodermi
└─┬─Chondrichthyes
└─Teleostomi
|
Contents
Overview |
|
| Lanarkia: a thelodont, showing the flattened, unarmoured body typical of this group. Graphic from Faktaside om fossile urfisk fra Norge, © 1998 Paleontologisk museum - University of Oslo. |
We treat the Thelodonti as a very large clade including osteostracans, gealeaspids, gnathostomes, and other forms, as well as the classical thelodonts. We do not mean in the process to forget the traditional thelodonts -- small, jawless fish with thelodont scales.
These "classical" thelodonts differ from the Cephalaspidomorphi (i.e. the armored jawless fishes) chiefly in lacking armour plates. Their bodies were covered entirely in tiny placoid-like scales. The scales of thelodonts are small (usually 0.5-1.5 mm) and robust, unlike the large flat scales of anaspids. Thelodont scales are conspicuous in having a dentine-covered crown, sometimes elaborately ornamented, and a bony base with a large central pulp cavity. The base has historically been regarded as "growing," because of the presence of Sharpey's fibers. The scales vary in form and size from the orbit, head, gill-covers, fins and trunk areas. Long (1993).
The thelodont fishes range in time from the Middle Ordovician - to the Frasnian Age of the Late Devonian. These scales frequently occur as microfossils and are widespread in the Late Silurian and Early Devonian, known from all the biogeographic provinces, although by the Middle and Late Devonian they appear to have become restricted to Gondwana Australia, Antarctica, Iran). The only known Late Devonian (Frasnian) thelodonts come from Western Australia They are an important group biostratigraphically, known principally from isolated scales, especially during the Early and Middle Devonian. Thelodont scales have been found in facies from all environments - marine, marginal marine and continental (fresh-water).
It used to be believed that thelodonts were a distinct group or clade of Pteraspidomorphs, related to the Heterostraci but distinguished by their lack of head shields and other features. Many workers now believe that they are an artificial group representing several different lineages that had lost or -- perhaps more likely -- failed to develop, armour. This is still a somewhat controversial point. However, as Janvier & Blieck (1993) note:
There is no clear evidence of the monophyly of the thelodonts, as such a micromeric exoskeleton is just what can be expected in primitive, generalized vertebrates. Only the peculiar 'handle' which prolongs anteriorly the scale base in most thelodonts may possibly be considered as a synapomorphy (unique feature). Thelodont scales display a wide range of diversity in morphology and histology, and their shapes vary within the individual animal. Their histologic structure varies from orthodentine (Turinia, Thelodus) to mesodentine (Phlebolepis), but their base is always made up of acellular bone (aspidin).
Only a few thelodonts are known from articulated specimens (Turinia, Loganellia; Phlebolepis), some of which allow rough reconstructions. The body shows well-developed paired lateral fins (presumably true pectoral fins), indicating good stability, dorsal and anal fins, and a heterocercal tail (the lower lobe of the tail is elongated). A typical Later Silurian genus like Lanarkia had a mouth located slightly ventrally (on the underside) on a flattened head, indicating possible bottom-dwelling habitats and that it fed on the seabed. A few types however, like the furcacaudiform thelodonts from the Early Devonian of Canada shown on the right, had the more typical fish-like body. Whatever their preferred environments, all thelodonts were clearly good swimmers. The cause of their downfall is not clear, but it may be that they were supplanted by the rise of true jawed fishes.
MAK. Revised ATW030621.
Thelodonts are the most enigmatic of all Paleozoic jawless vertebrates, because their internal skeleton was not ossified and their exoskeleton was made up of minutes scales which were often dispersed during decay.
Van der Brugghen & Janvier (1993)
Most thelodonts are not known from articulated specimens. We know them only from scattered scales. These can be found by the thousand in suitable Silurian and Devonian exposures; and they are the most common vertebrate microfossil in most Silurian sediments. Accordingly, thelodont scales, like conodont fossils, have attained a certain currency as stratigraphic markers for the Middle Paleozoic. But this currency is not too widely used. About two thirds of all thelodont papers published in the last decade bear the names of at least one of a tiny group of perhaps half a dozen active thelodont workers.
This combination of stratigraphic interest and a very limited pool of experts is unhealthy for the field. We intend no criticism of the practitioners of the arcane arts of thelodontology. (If nothing else, we would not attempt to pronounce, much less sully, the names of Drs. Tiiu Märss and Valentina Karatajūtē-Talimaa). The problem is simply one of purpose and numbers. Microfossil stratigraphy is not a phylogenetic or evolutionary study. It is a matter of correlating morphology over disparate samples. The stratigrapher is expected to recognize patterns, similarities and difference between scales from different locations, matrices, portions of the fish's body, and states of preservation. However, morphology is always in the eye of the beholder. When there are only a dozen eyes involved, no matter how experienced and objective those eyes may be, the chance that subjective or idiosyncratic factors have influenced the identification is very high. This is, if you will, an inherent diseconomy of small-scale science.
To make matters worse, the field lacks consistent terminology, even for such basic matters as the parts of the animal's body, or reliable definitions even for terms which everyone uses. How, for example does one distinguish between pulp depression, pulp cavity, and pulp canal? Can we be sure that the cap histology is orthodentine or mesodentine? What, exactly, is a "transitional scale"? There are no precise, testable definitions for these terms. Some have wondered, even aloud, whether these terms really have any meaning at all. They may describe vague regions of a continuum, or even completely subjective gestalt impressions. The history of vertebrate paleontology is itself littered with the bones of nomenclature which failed for one or both of those reasons.
With all that carping and kvetching said, we still have to deal with thelodont-craft as it actually is today.So we will turn to the scaled-up model of some model scales, below. These are thin sections, cut vertically, through the middle of the scale. We are looking at the scales in lateral view, with anterior to the right.
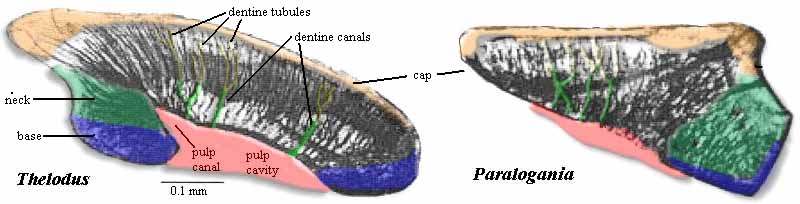
The first thing to notice is the base. The base is conventionally said to be aspidin, acellular bone. Frequently, it is relatively thin; but sometimes it forms a massive structure larger than the rest of the scale. This may be important, since it suggests possible close-packing of the scales in a manner similar to armored fishes. The base is frequently folded back on itself, almost enclosing the pulp cavity, so that only one (or sometimes more than one) pores connect the pulp to the rest of the universe. The Thelodus scale has a well-defined pulp cavity. This is the home of the living cells that sustained the scale. Part of the base is folded in to create a pulp canal In some scales, both structures are more elaborately developed. There may be several pulp canals. In Paralogania, there is only a vague pulp depression. Note that the boundary between the pulp depression and the body of the scale is likewise ill-defined in the central region. This is one characteristic of mesodentine and of the katoporid type of scale. Some workers classify the loganiids within the Katoporida for that (and other) reasons.
Both of these scales have some recognizable neck region. This is not always the case. Some scales have no neck, and some have very much better-developed necks between the base and the cap. Within the cap, we observe tracks left by the retreating front of odontoblasts. The larger tracks are dentine canals and the arbitrarily smaller branches are dentine tubules In the orderly orthodentine of the Thelodus scale, the canals rise almost at 90° to the line of the pulp cavity and branch terminally. In the mesodentine of Paralogania the odontoblasts are often trapped in spaces within the bone, the canals are wider, and they are not nearly so orderly. Thelodont papers frequently remark on whether or the dentine canals increase in diameter as they approach the pulp cavity. Finally, the cap itself is covered with a layer of dentine or, in some cases, a more hypermineralized tissue referred to as enameloid. The surface of the cap may be ornamented in a wide variety of ways, some of which are illustrated below.
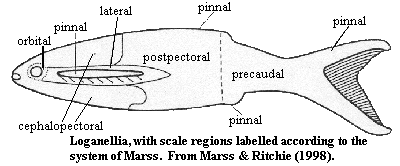
As we mentioned above, thelodont scales differ significantly depending on the part of the fish from which they are derived. There are two systems in use used to describe this. The traditional system, attributed to Gross, recognizes three rough categories: head ("oral" or "rostral"), "transitional," and trunk. This is generally adequate for most work on isolated scales -- one doesn't have a fish to work with, and it may not be necessary to describe variants in much detail. However, for more exacting work, it is sometimes useful to adopt the system of Marss 1986). This system is rarely used in its full rigor; but, for example, orbital scales are a distinct type in many thelodonts, and there is a rather marked difference between anterior and more posterior scales on the head.
Before developing that point, we should mention that this does not exhaust the catalog of scale types. Others frequently mentioned in the literature include:
It may be worth mentioning that the last category is in fact very minor. It is notoriously difficult to tell top from bottom in thelodonts. After well over a century, and numerous redescriptions, there is no consensus on whether the famous whole-body fossil of Turinia pagei is up or down. See review by Donoghue & Smith (2001) making compelling argument that it is belly-up). Similarly, there are perhaps a dozen good Scottish specimens of Lanarkia, but no one is quite sure whether the notochord continued into the lower part of the tail fin, as in most thelodonts, or into the upper lobe. Marss & Ritchie (1998).
During the course of her well-known study on the scales of Phlebolepis,Märss (1986) makes some interesting generalizations about scale positions, which go a long way towards imposing order on this mass of details, as well as coming close to reconciling the various systems of terminology in use at that time. The Roman numeral designations in the article have frequently been used by others, and are reproduced below:
These last three categories are referred to collectively as body scales, trunk scales, Rumpfschuppen, or Туловищные (= Туловищые?).
As Märss points out, there are parallels between this pattern and the squamation of living teleost fishes. The implication is interesting. Thelodonts would have had little need for this kind of hydrodynamic refinement unless they swam often enough to benefit from the gain in efficiency, and fast enough to generate the turbulent flows on which the efficiency gain depends. But, unlike modern fishes, most thelodont squamation, especially in the cephalopectoral region, was not imbricating. That is, the scales did not overlap so as to create a continuous surface. Thus, if thelodonts were nektonic, we might expect to see more dorsoventral asymmetry, since each scale could operate as a tiny hydrofoil, helping to provide lift. ATW030622
Salmo > Psammolepis. Obviously, our home-made definition.
From the Late Ordovician.
Vertebrata:::: Pteraspidomorphi + *: ((Nikoliviidae + Furcacaudiformes) + Thelodontida) + ((Katoporida + Cephalaspidomorphi) + (Loganiidae + Gnathostomata)).
Small (10-20 cm); usually (except furcacaudiforms, probably Shielia, and perhaps others) dorsoventrally compressed [DS01]; blunt rostrum [DS01]; mouth terminal [DS01]; probably no prenasal sinus distinct from nasopharyngeal cavity (possible $) [DS01]; orbits anterolateral [DS01]; orbits small [WC93]; primitively multiple gill openings; homocercal or hypocercal caudal fin [DS01]; many with paired pectoral fins without fin rays (which may or may not be homologous with gnathostome pectoral fins) [DS01$]; anal fin may be present [DS01]; probably all have lateral line pores enclosed in scales [M86]; small trilaminar scales with pulp cavity; sensory lines in canals, not grooves [DS01$]; scale base with Sharpey's fibers & assumed capable of growth [T99]; head scales usually rhombic, with shallow bases bearing vertical projections, and round to oval crowns with strong ridges arranged radially about the center [M86]; transitional scales oval to rhomboidal, with base shallow and/or protrudes anteriorly beyond crown, and with crown notched or ribbed anteriorly and often unornamented centrally [M86]; post-pectoral body scales often rhomboidal with central longitudinal ridge and several to many lateral ridges, the ornament often including spines [M86]; smooth transition between postpectoral & precaudal scales [M86]; precaudal (trunk) scales narrower & elongate oval to wedge-shaped, bearing ornament including sharp median ridge and fewer, but more pronounced, lateral ridges [M86]; caudal & paired fins with small, narrow, cuneiform scales with pointed posterior apex, ornamented with high-relief ribs and grooves tending to cover the entire crown [M86]; mostly open shelf and some near-shore environments [T99].
Many of the morphological characters detailed above do not apply to the possibly unique furcacaudiforms [DS01].
Thelodont scales are the most common vertebrate remains in the Early and Middle Silurian [T99].
This is inferred from the fact that whole animal fossils are normally found in dorsal or ventral orientation, with the (presumably laterally compressed) tail twisted to one side. [J96].
The association of a non-growing crown with non-growing base may be a synapomorphy [J96$].
Marss [M86] notes that Cenozoic fishes also have cycloid scales, without spines, anteriorly, and ctenoid scales, with longitudinal patterns, posteriorly.
Donoghue & Smith (2001) [DS01]; Janvier (1996) [J96]; Marss (1986) [M86]; Turner (1999) [T99]; Wilson & Caldwell (1993) [WC93].
ATW030615
Amaltheolepis, Gompsolepis, Nikolivia
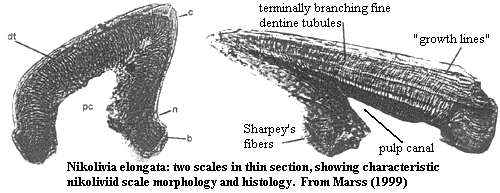
Thelodonti::: Furcacaudiformes + *.
Middle Silurian to Early Devonian (Lochkovian) of Arctic Canada, North America, Spitsbergen, Baltica & Russia
base small, low & anterior [BG02]; basal layer relatively thin [BG02]; pulp cavity large, with pulp canal [BG02]; dentine canals long & narrow [BG02]; dentine tubules branched [BG02]; crown large [BG02].
Blom & Goujet 2002) [BG02]; Marss (1999) [M99].
ATW060126.
Canonia, Cometicercus, Drepanolepis, Furcacauda, Pezopallichthys, Sphenonectris. The goldfish of the Paleozoic.

Middle Silurian to Early Devonian of Arctic Canada [WC98] & Siberia [K02].
Thelodonti::: Nikoliviidae + *.
Laterally-compressed, deep-bodied, fork-tailed forms [WC98]; mouth small and terminal [WC98]; eyes lateral and immediately anterior to branchial openings [WC98]; eyes large [WC93]; oralobranchial opening may have expanded ventrally in life [WC98]; branchial openings in a slanted, curved row [WC98]; branchial openings separated by rows of very small, composite scales joined at base, as in Loganellia [WC93]; hump-backed appearance [WC98]; large barrel-shaped stomach, tending to sharply rectangular appearance in endocasts [WC98]; short, straight intestine to anal notch [WC98]; caudal peduncle short and dorsoventrally deep [WC98]; caudal fin with long dorsal and ventral lobes and 8-14 intermediate lobes [WC98]; possible scaled connecting web in Devonian forms [WC93]; dorsal fin small or absent [WC98]; prominent paired lateral fins often present extending from base of gill row [WC98]; possible indications of suprabranchial paired fins or flaps in some forms [WC98]; thelodont scales usually < 400µ long, with large central pulp cavity [WC98]; orthodentine? [K02]; scales mostly of nikoliviid or loganiid type [WC93]; distinct base, neck & crown [WC98]; sensory canals probably present [WC98]; probable determinate growth [WC93]; deep water environment [WC98].
Some members of this group were originally described as dorsoventrally flattened forms, perhaps a thelodont version of an Osteostracan. As described and reconstructed by [WC98], the furcacaudiforms are more like thelodont goldfish. This configuration places the rather large orbits in a direct line between the gill slits and the small, terminal mouth. From a functional standpoint, this is downright weird. It also requires the brain to be oriented in a peculiar manner, and leaves no room at all for an olfactory chamber. There is also the matter of the "stomach endocasts" which have a strange, sharply rectangular appearance. However, the laterally flattened interpretation accounts nicely for the structure of the characteristic heterostracan-like tail, which would otherwise be almost impossible to explain.
Untitled Document; Les nouvelles (French); The oldest stomach in the world; Furcacaudiformes; The Journal of Vertebrate Paleontology abstract of [WC98]); Lower Devonian (Lochkovian) thelodonts from October Revolution ...; EXN.ca | Discovery.
Karatajute-Talimaa 2002) [K02]; Wilson & Caldwell (1993) [WC93]; Wilson & Caldwell (1998) [WC98].
ATW030225.
checked ATW050524