Cephalaspidomorphi: Osteostraci
Pituriaspids plus Ateleaspis, Hirella and Hemicyclaspis
Taxa on This Page
- Ateleaspis X
- Hemicyclaspis X
- Hirella X
- Osteostraci X
- Pituriaspida X
Pituriaspida
 Pituriaspida was erected in 1991 based on armours of two genera (Pituraspis and Neeyambaspis) of rostrate (nose spiked) jawless fishes known as yet from a single locality of sandstones in central Australia (Georgina Basin, Western Queensland). Pituraspis illustrated here) has an elongate headshield (with an area of attachment for paired fins, as in the Osteostraci), Neeyambaspis a short triangular one. The exoskeleton is ornamented with tiny rounded tubercles. The outline of the rest of the body shown here is pure fancy. Almost nothing is known of the rest of the body, apart from the headshield, and even that is poorly preserved. These creatures are very tentatively placed close to the Osteostraci on the phylogenetic tree. MAK001204.
Pituriaspida was erected in 1991 based on armours of two genera (Pituraspis and Neeyambaspis) of rostrate (nose spiked) jawless fishes known as yet from a single locality of sandstones in central Australia (Georgina Basin, Western Queensland). Pituraspis illustrated here) has an elongate headshield (with an area of attachment for paired fins, as in the Osteostraci), Neeyambaspis a short triangular one. The exoskeleton is ornamented with tiny rounded tubercles. The outline of the rest of the body shown here is pure fancy. Almost nothing is known of the rest of the body, apart from the headshield, and even that is poorly preserved. These creatures are very tentatively placed close to the Osteostraci on the phylogenetic tree. MAK001204.
The Osteostraci (also called the cephalaspids, meaning 'head shields') are an important group of Silurian and Devonian fishes from the Euramerican province. They were flattened bottom-dwellers (triangular in cross-section) that sucked up food particles from the seabed through a rounded mouth on the underside of the head. Despite their bottom-dwelling habits, they were good swimmers. Unlike many other jawless fishes, some osteostracans possessed paired pectoral (front) fins, a dorsal fin on the back near the tail, and a strong epicercal (upturned) tail. The enlarged upper lobe of the tail produced lift at the rear of the body, keeping the animal's head down while it sucked up food from the seabed. These were possibly active animals and good swimmers, despite their heavy armour.
The large and usually horseshoe-shaped head shield (cephalic shield) was made of a single plate of undivided bone. There are many variations in the shape of the cephalic shield, from simple, semi-circular elongate oval shaped forms. In most forms the corners of the head shield were drawn out into keel-like cutwaters, the backwards protruding cornuta or "horns", e.g. Cephalaspis, Others forms, like Boreaspis, have an anterior rostrum ("nose" spike).
 Since the head-shield was made of one piece of bone, it is unlikely that the shield grew during the animal's adult life, unlike the individual bony head plates of the heterostracans. It is thought that osteostracans had an unarmored larva, and that the bony shield developed only when the creature was full-grown.
Since the head-shield was made of one piece of bone, it is unlikely that the shield grew during the animal's adult life, unlike the individual bony head plates of the heterostracans. It is thought that osteostracans had an unarmored larva, and that the bony shield developed only when the creature was full-grown.
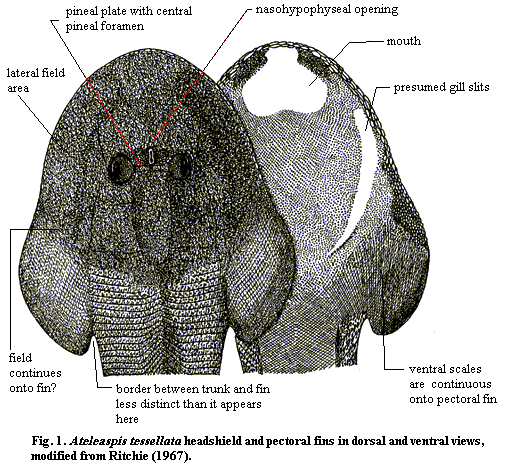
Another unique characteristic of this group is presence of concentrated patches ("fields") of sensory organs developed on both sides, and on top, of the head shield (referred to as the lateral (front) and median or dorsal behind the eyes) fields). These are depressions covered with dermal platelets and connected by canals to the otic capsule. These organs were richly supplied with nerves (see left), and must have detected water-borne vibrations. Alternatively, they could have been electric organs. The "fields" have been variously interpreted. They were most certainly of a sensory nature (perhaps generating a mild electric field that allowed the animal to navigate in murky water). The headshield also has two separate eye holes, a pineal opening within a small pineal plate the eyes and a key-hole shaped nasohypophysial opening in front of the eye holes.
The anatomy of osteostracans is well known because of a unique development among these jawless fishes. Bone was laid down inside the body, in a thin layer over the cartilage of the skeleton. From this fossilized bone, the detailed structure of the brain, gills, mouth and even individual nerves and blood vessels can be traced. The brain (above) appears to be similar to that of modern lampreys.
The body is covered with large scales. Both the endo- internal) and exoskeleton is made up of true bone with cell spaces (the heterostracans have acellular bone). The Osteostraci share a number of unique characteristics - synapomorphies - with the gnathostomes or jawed fish true bone, epicercal tail, sclerotic ring, endolymphatic openings, dorsal jugular vein, concentrated paired fins) and were clearly closely related.
The Osteostraci are biogeographically limited to the Euramerican Province (Laurentia and Baltica), and are useful biostratigraphic indicators. They are known from the Late Wenlock (Middle Silurian) to the Frasnian (Late Devonian), but were at the height during the period of the Late Silurian and Early Devonian. Strangely, the earliest (Wenlock) occurrences are represented by both very primitive (Ateleaspis, Aceraspis) and also very advanced (Tremataspididae) members of the group. This indicates either that the diversification of the group took place before the earliest known occurrences, or that the presently accepted phylogeny is completely wrong. Osteostracans are rare in the Devonian of North America (West Euramerica) and practically none of the North American taxa can be assigned with certainty to any of the major European forms.
Most osteostracans belong to a large monophyletic group, the Cornuata, characterized by pointed cornual processes in front of paired fins. The Cornuata comprise five major monophyletic groups, and a few minor groups of uncertain affinities. There are also a few genera of non-cornuate osteostracans which probably represent the primitive osteostracan condition (Ateleaspis, Aceraspis, Hirella, Hemicyclaspis).
Osteostraci: Taking arms against a sea?
The more one studies the Osteostraci, the less one knows with any degree of confidence. The conventional wisdom -- which, right or wrong, is frequently all the wisdom there is -- states that the Osteostraci are the "most advanced agnathans on the evolutionary ladder leading to higher vertebrates." Long 1995). That is, they are jawless fish, but with ossified sclera, paired pectoral appendages (presumably for gripping ladders) and other characters which unite them with gnathostomes. The basic wiring of the brain is fairly well known due to the unique architecture of the skull and Stensiö's destructive thin-sectioning of irreplaceable specimens, an incredible gamble that no one would now be permitted to take, but which paid off remarkably well. The brain turns out to be quite similar to the lamprey brain, which is also taken as a sign that the Osteostraci are but a fruit stand on the broad highway to the gnathostomes.
This general take on the Osteostraci is not disputed, least of all here. The true purpose of this essay, to the extent it has one, is to express frustration with the details of these osteostracan pectoral appendages. They are probably homologous with gnathostome pectorals, but there are problems.
The first problem is stratigraphic. Bits and pieces believed to be gnathostome, specifically chondrichthyan, are known as early as the Late Ordovician. Osteostracans are not known before the Early Silurian. Of course paleontology is full of these little stratigraphic anomalies, and they often mean nothing. However, consider that the fossil record of osteostracans is quite good. They have a lot of hard parts, tend to hang around in silty, low-energy marine environments, and have generally shown themselves to be taphonomically friendly to scientists. By contrast, chondrichthyans are an unruly mob with few calcified parts. They often wander about aimlessly in pelagic environments, and have repeatedly demonstrated a willful, even blatant, indifference to the legitimate needs of hominid paleontologists. Under these circumstances, the stratigraphic problem becomes more significant. If the gnathostomes came first, the osteostracan pectoral may be convergent, and not homologous.
The second problem is that the pectoral appendage does not seem to have been a great success among the Osteostraci. In many cases, it not certain that the appendages existed at all. The structure is certainly never elaborated or developed. In osteostracans with elongated shields, the paired limbs could not have existed at all; and no osteostracan shows any sign of paired pelvic fins. To the contrary, the more advanced Osteostraci are cornuates. These osteostracans developed enormously broad head shields, very much like those of a number of galeaspids, with long posteriorly-directed processes at the edges. In these forms, it is difficult to see how the pectoral fin (if it existed) would have been of much use in navigation. Indeed, since the Osteostraci seem to have had only two semi-circular canals, it is difficult to see how anything could have been of much help in navigation since the fish evidently lacked the neural wiring to orient themselves well for movement in a three-dimensional medium.
The third problem is structural. The osteostracan pectoral fin is directed antero-posteriorly. That is, it is expressed as a posterior process from the back of the skull, not a lateral process of the trunk. That is not too unexpected, since the pectoral fin girdles are essentially part of the skull even in teleosts. However, the osteostracan paired fin has no well-defined girdle, no fin rays, and no endoskeleton. There are what appear to be attachments for muscles, some scale rows which may have added rigidity, and certainly the fin was integrated with the circulatory system. Still, the appendage is supplied by the branchial artery, not a separate subclavian. The case could perhaps be made that we are looking at the base of a flexible external gill, rather than a limb. The position of the appendage puts it in the hydrodynamic "shadow" of the large head shield. Not a bad place to put fragile external gills, but surely an odd location for control surfaces, and the orientation is entirely different from the pectoral fins of living gnathostomes.
For that matter, the vertebrate pectoral limbs may simply have been external gill supports which were exapted for mobility. One of the leading theories on the subject is not too distant from that view. Still, it seems a long leap from these rather formless flaps of presumably ectodermal tissue to the fins of extant gnathostomes, which seem to be developmentally driven by their mesodermal components. The conventional wisdom is probably quite correct, but there remains room for serious doubt.
Ateleaspis: Fins and Fields
 Ateleaspis has one notable distinction for which it deserves more than casual interest. It is the most basal craniate to have unmistakable paired appendages. The pectoral fins of this genus are not terribly impressive. They lack any discernible skeletal elements. They may not even be homologous (in the phylogenetic sense) with arms. However, there is absolutely no doubt that they exist. The same cannot be said for the often-cited, but rather uninformative, lateral fin folds of anaspids. For more details, see Anaspida.
Ateleaspis has one notable distinction for which it deserves more than casual interest. It is the most basal craniate to have unmistakable paired appendages. The pectoral fins of this genus are not terribly impressive. They lack any discernible skeletal elements. They may not even be homologous (in the phylogenetic sense) with arms. However, there is absolutely no doubt that they exist. The same cannot be said for the often-cited, but rather uninformative, lateral fin folds of anaspids. For more details, see Anaspida.
A second curiosity of Ateleaspis, and of most osteostracans, is the presence of sensory fields of unknown function, usually located along the mid-line of the headshield, behind the orbits, and along the lateral margins of the headshield. As mentioned in the main entry. These "fields" are believed to be made up of loose platelets of bone connected to a complex of bifurcating canals, nerve channels, endolymphatic ducts, or whatever. These channels eventually connect with the brain within the labyrinth.
Possibly, these two features are related. Let's review what we know about the fields. They are made up of small platelets which, in the basal Ateleaspis, are not at all different from the surrounding dermal bone of the headshield. In more derived species, they become more distinct and specialized. However, they seem to have been able to accomplish their essential function as simply generalized bits of dermal bone. These platelets lay over a regular, bifurcating system of channels connected to the labyrinth -- the organs which detect orientation in a gravity field, or acceleration (relativisticly the same things). The Osteostraci, like other pregnathostome craniates, had only two pairs of semicircular canals in the labyrinth. That is, their kinesthetic sense was somewhat handicapped in dealing with fully three-dimensional movement. Presumably Osteostraci were able to sense roll and yaw: rotation around an antero-posterior axis and around a dorso-ventral axis, respectively. Since it is impossible to imagine a vertebrate without the ability to detect roll or yaw, it is generally believed that what was missing or deficient was a sense of "pitch," rotation around a horizontal axis, corresponding to motion up or down in the water column.
But, while the Osteostraci lacked a third semicircular canal, they possessed the fields, which were also connected to the labyrinth. The field platelets in Ateleaspis look just like the rest of the shield. They aren't specialized sensors. All that happens is that, when the fish moves "uphill," the platelets are pressed down a bit harder on the channels below them. This sends some kind of signal, pressure wave or electrical, directly to the portion of the brain which computes orientation and movement in space. We can be fairly confident in this conclusion because the effect of the mechanical pressure of a small plate on either neural tissue or a fluid-filled duct would be to send a signal to the brain -- whether or not this were the "intended" effect. The same, simple system would register a different types and patterns of signal for turbulence caused by currents, waves, or other fish.
This looks well enough, but why bother with this clumsy arrangement when Ateleaspis had a perfectly good lateral line system, and perhaps even electrical senses? The neuromast organs of the lateral line are similar (and closely related to) the receptors in the labyrinth. In modern fish, they are sensitive to very slight water currents. Assuming the osteostracan lateral line is not too different, where is there a need for a gross mechanical receptor of this sort?
There is no simple answer. However, there is also no good evidence that osteostracans actually had lateral line organs except on the head shield. Some, like Ateleaspis, had no ventral lateral lines, even on the head. Perhaps their lateral line system was not as effective as in Recent fish. As well, lateral lines are not constructed in a manner which lends itself to detecting flow normal to (towards or away from) the surface -- loosely speaking, movement up and down -- or to making sense of water movements which have both local turbulence and large-scale flow. So, perhaps there was room for a sense which was strong where the lateral line was less dependable.
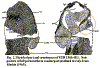 Whatever the sensory fields do precisely, it may have to do with the pectoral limbs. There is a fairly striking correlation between the extent of the sensory fields and the extent of the pectoral fins. In many cases, the fins are not actually known. However, the extent of the field is fairly closely correlated with the relative size of the fins (where known).
Whatever the sensory fields do precisely, it may have to do with the pectoral limbs. There is a fairly striking correlation between the extent of the sensory fields and the extent of the pectoral fins. In many cases, the fins are not actually known. However, the extent of the field is fairly closely correlated with the relative size of the fins (where known).
In the primitive Ateleaspis, the fields appear to actually extend onto the fins. Although it may be a taphonomic artifact, one of the specimens described by Ritchie suggests that the pectoral fin extends well beyond the relatively scaleless dorsal area and is actually limited anteromedially only by the line of the gill arches. The field area of the fin is indicated on the sketch. As Ritchie states, the fin is plainly an extension of the headshield. It is not a lateral breach in, or extension of, the body wall in the manner of modern tetrapods. Thus, the fields and the paired limbs are not only correlated; they are physically associated at least in this basal species.
If the foregoing chain of speculation (one could not call it "reasoning") has any validity, the most probable interpretation is that the fins and fields developed together: a means of stabilizing swimming along with the sense organs to provide feed-back control for it. This reconstruction has the virtue of elegance and economy, but it may suggest that the osteostracan pectoral limb is not strictly homologous with the gnathostome limb. However, this a suggestion built on a speculation -- going a bit too far out on a limb, so to speak. ATW 001016.
Descriptions
Pituriaspida: Neeyambaspis, Pituriaspis.
Probable paired pectoral appendages. Similar to Galeaspida. The only unique characteristic of the Pituriaspida is a peculiar pit, of unknown function, ventral to the orbits; marine or deltaic environment.
Links: Pituriaspida; 116.htm; DEVONIANO peixes agnatas. ATW020615.
 Osteostraci: = Cephalaspididae):
Osteostraci: = Cephalaspididae):
Range: Early Silurian to Late Devonian of North America, Europe and Russia.
Armored jawless fish (5-100 cm) with massive cartilaginous skulls and paired pectoral fins. Extensive cephalic endoskeleton of cartilage covered with perichondral bone; large cephalic plates with "cephalic fields" (shallow dorsal depressions covered with small polygonal scales and connected to labyrinth cavity by canals); 2 semi-circular canals; mouth and up to 10 pairs of branchial openings ventral; probably with velum-like pumping organ; orbits close to dorsal midline, with sclerotic ring and commonly ossified sclera; central nasal opening and small pineal opening; cornua common; some with dorsal spines or ridges on head shield or elongated shields; heart in chamber posterior to skull, probably 2-chambered, with ventricle and atrium lying side by side; some details of circulatory system known; ossified dorsal arcualia on notochord; 1-2 dorsal fins or scale ridge; also paired lateral scale ridges; heterocercal tail; frequently small horizontal lobe below caudal modified anal fin?); many have vertical rows of squarish scales on caudal fin; paired fins present as posterior outgrowths from base of broad head shield; pectoral fins with complex musculature but no known fin rays or endoskeleton; pectorals may be stenobasal or eurybasal; from low energy near-shore or estuarine marine sediments; exoskeleton with cellular bone and subaponevrotic circulation; make-up of exoskeleton highly complex and variable, normally with mesodentine outer layer for more detail, see note under Benneviaspidida); lateral line system on trunk; ventral surface of small plates.
 Ateleaspis: = Aceraspis):
Ateleaspis: = Aceraspis):
Phylogeny: Osteostraci: (Hirella + (Hemicyclaspis + Cornuata)) + *.
Primitive, 15-20 cm osteostracan; the most basal known vertebrate with paired fins. Headshield of cellular bone, heavily ornamented with dentine tubercles; tubercle size increases near orbit and headshield margins; headshield flattened, with a broad, flat basal margin containing "fields" (areas made up of loosely attached small plates over poorly understood bifurcating canals connected to the labyrinth area of the brain); endolymphatic duct opens inside a median "field" running along raised peak of posterior headshield; "fields" are larger, but less distinct less specialized?) than in more derived osteostracans; orbits have ossified sclera as incomplete hollow sphere; orbits closely set and joined by a rod-like pineal plate; pineal foramen runs through center of pineal plate; keyhole shaped nasohypophyseal opening in a "heart-shaped" depression just anterior to pineal plate; poorly-defined ventral mouth; ventral surface of head covered with uniform, minute scales; gill openings along lateral margins of ventral surface; sensory line system present on headshield, perhaps on trunk; cornuate headshield processes absent; cranial endoskeleton poorly developed; two distinct dorsal fins without median dorsal ridge scales; 2nd dorsal and caudal fins have posterior fin membrane; trunk has narrow basal marginal flange; broad-based pectoral fins; gradual transition from both tesserate "field" of headshield margin and brick-like scales of trunk and flange; pectoral girdle and rays (if any) unknown; pectorals appear to be joined to trunk or headshield along most of their length; pectoral appear as extension of lateral headshield. Likely inhabited low-energy marine or perhaps estuarine areas; found as a rare species in association with eurypterids, anaspids & thelodonts.
Links: Osteostraci; Biology 356; Biology 356; The exoskeleton of Ungulaspis and Ateleaspis (abstract, histology).
References: Janvier 1996); Ritchie (1967). ATW031130.
Hirella:
Characters: ventral platelets on headshield; pectoral fin base narrow. ATW000624.
 Hemicyclaspis: H. murchisoni
Hemicyclaspis: H. murchisoni
Range: Late Silurian to Early Devonian of North America & Europe.
Characters: narrowed, more differentiated sensory fields; anterior dorsal fin reduced to large median scute.
Links: Dinosaurs: Model & Collectible Pricelist; Fish Replicas - Hanman's Fossils & Minerals; geologic; Volume 16 (V2) (small, badly drawn, but accurate sketch); 120.htm; museum für naturkunde berlin: glossar geschichte der wirbeltiere (German); view.cgi?page=2&corner=18&file=29 Japanese); Vertebrate Korean); Världshaven på Unga Fakta! Swedish); Genomgång Ostracoderma Swedish); 古生物学研究的意义 Japanese); おもちゃのサ・カ・ナ models); DEVONIANO peixes agnatas; 척추동물아문(Vertebrate, 등뼈를 가진다) Korean: notes on feeding style?); 展示室にある標本 Japanese: nice model); The Devonian Period of the Paleozoic Era.
References: Janvier (1996). ATW031022.