
The secret of conodont survival: Evolutionary fitness or corporate sponsorship?
(with apologies to Albanesi & Barnes 2000)
| Conodonta | ||
| The Vertebrates | Overview |
| Vertebrates Home | Vertebrate | Vertebrate |
|
Abbreviated Dendrogram
Vertebrata
├─┬─Pteraspidomorphi
│ └─Gnathostomata
└─Conodonta
├─Paraconodontida
└─Euconodonta
├─Proconodontidae
└─┬─Protopanderodontida
└─Prioniodontida
├─Balognathidae
└─┬─Prioniodinida
└─Ozarkodinida
|
Contents
Index |
| "We specifically reject terms such as 'conodont- bearing animal' or 'conodont animal' -- at least until our colleagues who study brachiopods or dinosaurs adopt the rubrics of 'brachiopod animal' or 'dinosaur animal' to distinguish between the shells or bones of those creatures and the entire organism." Sweet & Donoghue (2001) |
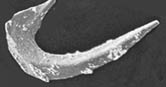
One of the true, great mysteries of vertebrate paleontology concerns the Conodonta, and the relation of the conodont apparatus to vertebrate bones. Indeed, until a decade ago, these matters were sufficiently mysterious that very few suspected a relationship to vertebrate paleontology at all. The very first paper published on conodonts actually had reported them as the remains of some unknown fish. Pander (1856), vide Sweet & Donoghue 2001). But this notion was quickly discarded. It is really only since the mid- 1990's that Pander's original intuition has been proved correct and these weird Paleozoic worms have been fully initiated into the exclusive sorority of extinct vertebrates. See, e.g., Janvier (1995), Donoghue et al. (2000).
The reason for this long-delayed acceptance into polite society relates -- as it sometimes does -- to the fact that conodonts are entirely too common. Conodont elements, the disarticulated bony remains of the complex conodont oral apparatus, are found as microfossils by the tens of thousands in virtually every Paleozoic marine sediment, everywhere in the world. Further, the various elements occur as isolated pieces, mixed more or less randomly in matrix. With respect to each, as Shelley 1817) noted of another disarticulated relic, "nothing beside remains. Round the decay ... boundless and bare, the lone and level sands stretch far away" It was not until the 1960's that a few, spatially contiguous remains were recognized as "natural assemblages," collections of conodont elements which represented the parts of a complex but unified structure belonging to a single organism which had remained in situ, where the conodont had died. With these discoveries came the whole notion of a "conodont apparatus," a unique food-processing machine made up of conodont elements, and the search for the correct three dimensional arrangement and functional biology of the elements was under way.
|
|
|
|
The secret of conodont survival: Evolutionary fitness or corporate sponsorship? |
||
Although the reconstruction of the apparatus has been a central theme of conodont research in recent years, we will defer any serious discussion of the apparatus until we are positioned to look at the specifics of the Ozarkodinida and the work of Purnell & Donoghue (1997) on this, probably the best-known group of conodonts. Instead, we will take up the very poorly known matter of conodont biology and chordate affinity.
As an aside, conodont biology has been the source of one of the strangest social transformations in recent academic history. Consider that, for many years, conodont affinities were unknown, even as to Kingdom, much less phylum. However, conodont elements are very common items and easy to isolate. As a result, conodont elements became key stratigraphic markers. The students of these obscure phosphatic scraps were thus much in demand as petroleum stratigraphers. While stratigraphy is a relatively low academic calling, it pays rather well because of its obvious real-world commercial value. By contrast, vertebrate paleontology has always been a useless, but popular and respected pursuit, although the pay ranges from the execrable to the non-existent. Imagine, then, the dismay of all concerned when conodonts were discovered to be vertebrates. Suddenly, conodont workers were dragged from the oil-stained proletarian legions of Houston and Riyadh petro-imperialism and thrust into the rarified, if ragged, intellectual circles of vertebrate paleontologists.
But we must retreat from these sociological speculations to return to the more pertinent, if scarcely less speculative, matter of conodont biology. As noted previously, a very few conodont fossils are preserved as natural assemblages. Of these rare events, a scant handful may also preserve traces of soft tissues on careful, or imaginative, inspection. From these, exceedingly unusual, fossils, conodont workers have recently begun to reconstruct the vertebrate inheritance of these chordate Cinderellas.
 A life reconstruction from Sweet & Donoghue (2001) is shown at right. These authors have recently reviewed the evidence for the chordate nature of conodonts. There would be little point in recapitulating this excellent and very readable review. However, we shall do so anyway on the theory that, if the reader had ready access to such materials, he or she would not be spending valuable time on this web site in the first instance. However, unlike Sweet & Donoghue, we will make no effort to strike a linguistically neutral pose, and will use vertebrate anatomical vocabulary without excuse or, perhaps, justification.
A life reconstruction from Sweet & Donoghue (2001) is shown at right. These authors have recently reviewed the evidence for the chordate nature of conodonts. There would be little point in recapitulating this excellent and very readable review. However, we shall do so anyway on the theory that, if the reader had ready access to such materials, he or she would not be spending valuable time on this web site in the first instance. However, unlike Sweet & Donoghue, we will make no effort to strike a linguistically neutral pose, and will use vertebrate anatomical vocabulary without excuse or, perhaps, justification.
Conodonts are elongate worm-like organisms that flourished from the Middle Cambrian through the end-Permian event. After near extinction in that catastrophe, they made a considerable recovery, but dwindled and disappeared by the end of the Triassic or perhaps the earliest Early Jurassic. Their phylogeny is very poorly known . Early Cambrian "protoconodonts" are currently believed to be the unrelated remains of chaetognaths or "arrow worms"). Sweet's (1988) pre-cladistic taxonomy is still the standard and has been adopted here. As of this writing (020727), a full-scale cladistic study is nearing completion and may see daylight in another two years.
 Much of the discussion of other cranial structures is based on a single specimen of Clydagnathus windsorensis, which is reproduced at right, with structures identified according to the interpretation of Donoghue et al. 2000), Sweet & Donoghue 2001), and concordant sources. Conodonts have a reasonably well-defined head with two large hemispherical or ovoid anterolateral structures which have been interpreted as eyes. For reasons which will be discussed presently, we believe this interpretation is incorrect. Posteromedial to the eyes are a pair of dark, circular structures identified as otic capsules. Posterior and slightly lateral to these are a series of paired vacuities which are interpreted as pharyngeal pouches.
Much of the discussion of other cranial structures is based on a single specimen of Clydagnathus windsorensis, which is reproduced at right, with structures identified according to the interpretation of Donoghue et al. 2000), Sweet & Donoghue 2001), and concordant sources. Conodonts have a reasonably well-defined head with two large hemispherical or ovoid anterolateral structures which have been interpreted as eyes. For reasons which will be discussed presently, we believe this interpretation is incorrect. Posteromedial to the eyes are a pair of dark, circular structures identified as otic capsules. Posterior and slightly lateral to these are a series of paired vacuities which are interpreted as pharyngeal pouches.
Far less debatably, the long conodont body has obvious V-shaped myomeres throughout its length. A pair of (very probably) dorsal parallel lines also extend the length of the body, and these are believed to represent the outlines of the notochord. Cartilaginous arcualia seem to be associated with the notochord. Last, and most convincingly of all, conodonts bear a distinct tail with a small but clear set of fins supported by very fish-like fin radials.
From this rapid tour of the principle scenic attractions of the taxon, it is reasonably clear that the Conodonta are chordates, and probably vertebrates. The only competing hypothesis -- chaetognath affinity -- provides some alternative interpretations; but the dorsal notochord, tail, and fin radials make this unlikely. For the present, despite many difficulties of detail, we can accept the hypothesis of vertebrate ancestry with a fair degree of confidence. ATW020727.
At this point, the reader who has still not been induced to depart these precincts in haste, or to resort to some more learnèd and authoritative source, is confronted with a small number of equally repugnant options. They are as follows:
1. Learn why the conodont eye is nothing of the sort. continues this discussion)
2. Ignore this baseless prattle and proceed to the phylogenetic summary.
3. Skip over to the Ozarkodinida for additional rumor and speculation concerning the conodont apparatus.
checked ATW031116