The Olenekian
The Olenekian Age of the Early Triassic Epoch: 245 to 250 million years ago
The Limits of the Olenekian
 The Olenekian Age is named for the Олениок River in Siberia, where the type section occurs, and which we have probably spelled incorrectly. If so, it would not be the first muddled fact about this age, which has gone by an embarrassing number of names, particularly for such a modest span of time. The first half of the Olenekian has been referred to as the Late Nammalian, Smithian, or Ayaxian. The later Olenekian is (more or less) the Spathian or Russian. The entire Age is the same as the Late Scythian. Internationally recognized correlations have only been worked out in the last few years, and it appears that there is still much uncertainty, particularly for regions far from the type locality -- which, unfortunately, includes most of the world.
The Olenekian Age is named for the Олениок River in Siberia, where the type section occurs, and which we have probably spelled incorrectly. If so, it would not be the first muddled fact about this age, which has gone by an embarrassing number of names, particularly for such a modest span of time. The first half of the Olenekian has been referred to as the Late Nammalian, Smithian, or Ayaxian. The later Olenekian is (more or less) the Spathian or Russian. The entire Age is the same as the Late Scythian. Internationally recognized correlations have only been worked out in the last few years, and it appears that there is still much uncertainty, particularly for regions far from the type locality -- which, unfortunately, includes most of the world.
The beginning of the Olenekian (Induan - Olenekian boundary) is marked by the first appearance of the conodont Neospathodus waageni. A great deal of Olenekian stratigraphy has historically been done using ammonoid biozones. The opening of the Oleneckian is also associated with the first appearance of a number of ammonoids, including Flemingites and Euflemingites. It occurs just before the end of the second Triassic normal magnetostratigraphic zone, and the peak of the first Triassic positive excursion of carbon isotope 13C. The end of the Olenekian, i.e. the Olenekian - Anisian boundary, is characterized by conodont zonation: the last occurrence of Neospathodus homeri and N. symmetricus and the first occurrence of N. gondelloides, Chiosella timorensis and Neogondolella regalis. Another indicator is a magnetostratigraphic shift from reversing polarity in the Late Olenekian to a long normal zone in the Early Anisian. Unfortunately, the absolute age of these terminal events is considerably less certain. It has been found, by various measures, to occur at anywhere from 240 to 247 Mya, with more recent measurements indicating older dates. As always, we have adopted the ICS/IUGS dates. ATW040206, rev'd ATW050429.
The Geography of the Olenekian

"Darkness Crumbles Away"[1]
 Many discussions of the end-Permian extinction go something like this: "the seas became thoroughly deoxygenated, and most marine Metazoans died off. On land, many species also died off, but a number of critical (to us) tetrapods survived unchanged. The die- off may have been caused by climate, comet, vulcanism, or virus." The discussion then moves on to the evolution of archosaurs in the Middle Triassic. What gets left out here is the thing that really separates the end- Permian from every other mass extinction of the Phanerozoic -- the incredibly slow rate of recovery. In fact, reasonably normal ecosystems are not seen again until the last half of the Olenekian, about six million years after the extinction event(s). Just as interesting is the trajectory of the recovery. For over five million of those years, the recovery proceeded at a snail's pace. Then, in the space of just half a million years, the process accelerated, and the world's living systems recovered something resembling their Late Permian vitality.
Many discussions of the end-Permian extinction go something like this: "the seas became thoroughly deoxygenated, and most marine Metazoans died off. On land, many species also died off, but a number of critical (to us) tetrapods survived unchanged. The die- off may have been caused by climate, comet, vulcanism, or virus." The discussion then moves on to the evolution of archosaurs in the Middle Triassic. What gets left out here is the thing that really separates the end- Permian from every other mass extinction of the Phanerozoic -- the incredibly slow rate of recovery. In fact, reasonably normal ecosystems are not seen again until the last half of the Olenekian, about six million years after the extinction event(s). Just as interesting is the trajectory of the recovery. For over five million of those years, the recovery proceeded at a snail's pace. Then, in the space of just half a million years, the process accelerated, and the world's living systems recovered something resembling their Late Permian vitality.
This peculiar sequence of events was documented for a large terrestrial ecosystem by Looy et al. 1999) who examined the palynological record of the forests of Europe. In the Lopingian Late Permian), Europe was covered in conifer forests. At the end-Permian, the conifers simply disappear. For the Induan and most of the Olenekian, the region was covered by Pleuromeia- dominated lycopsid stands. During this period, soil formation was low. Apparently the total biomass of lycopsids was not particularly impressive, or the rate of erosion was unusually high. Probably both factors played a part, as there were few tree- sized species, and the soils would not have been strongly held. This may well have been a world- wide phenomenon, in view of the global absence of Early Triassic coal deposits. Then, late in the Olenekian, the lycopsids were rapidly replaced by a transitional flora of low, shrub- like, and rather simple conifers. Within half a million years, this was succeeded by open, Voltzia-dominated conifer forests, not so different from the original cover of the Lopingian.
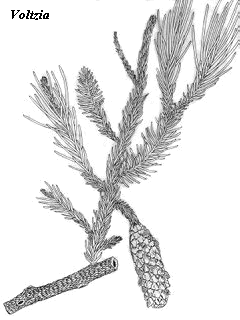 The authors point out that the succession resembles the recovery patterns after the Wisconsin glaciation at the beginning of the Holocene
-- but several thousand times slower. The comparison may be inexact. We do not yet know enough details of the Early Triassic recovery to attempt a rigorous comparison, and Holocene grasses are very unlike lycopsids. Still, the comparison is an interesting model. What could possibly have slowed a process which ought to have taken a few hundred years and stretched it out to five million? Climate is not the answer. Looy et al. point out that the same warm, semi- arid conditions are believed to have prevailed before, during and after the Early Triassic. They suggest that the appropriate conifers were all extinct and that tree- sized conifers simply needed time to re-evolve.
The authors point out that the succession resembles the recovery patterns after the Wisconsin glaciation at the beginning of the Holocene
-- but several thousand times slower. The comparison may be inexact. We do not yet know enough details of the Early Triassic recovery to attempt a rigorous comparison, and Holocene grasses are very unlike lycopsids. Still, the comparison is an interesting model. What could possibly have slowed a process which ought to have taken a few hundred years and stretched it out to five million? Climate is not the answer. Looy et al. point out that the same warm, semi- arid conditions are believed to have prevailed before, during and after the Early Triassic. They suggest that the appropriate conifers were all extinct and that tree- sized conifers simply needed time to re-evolve.
This explains too little. The Early Triassic recovery shows roughly the same trajectory everywhere -- probably. The uncertainties of Early Triassic stratigraphic correlation defeat any certain comparison. But marine invertebrates, temnospondyls, and tetrapods all show a recovery pattern similar to that of the pine forests. The timing might be similar because evolutionary events are somehow interrelated. However, there is no particular reason to think so. For example, East Asian radiolarian protists also show an extinction at the end-Permian, a depauperate population until the Late Olenekian, a Late Olenekian transitional community, and a characteristic Mesozoic community at the beginning of the Anisian. Suzuki et al. (2002). It's hard to think of a reason that Japanese radiolarian evolution would be correlated with European pine forests. As matters stand, there are no compelling explanations for the initial extinction, for the Earth's painfully slow convalescence, or for its much faster recuperation at the end of the Olenekian. ATW040207.
[1] "Darkness Crumbles away.
It is the same old druid Time as ever,
Only a live thing leaps my hand ..."
Break of Day in the Trenches
Isaac Rosenberg (1916)